Курсовая работа: Биогенез мембран
Курсовая работа: Биогенез мембран
Пензенский государственный педагогический университет
им. В.Г. Белинского
Курсовая работа на тему:
«Биогенез мембран»
Выполнил: студентка 4-го курса гр. БХ-4
Злобина Н.
Проверил: Соловьев В.Б.
Пенза, 2006
Оглавление
1 Введение
2 Образование и встраивание мембранных белков
2.1 Транспорт белков
2.2 Сигнальные последовательности белков
2.3 Стоп- сигналы переноса
2.4 Сигнальные пептидазы
2.5 Белки необходимые для распознавания сигналов переноса
3 Липиды
3.1 Синтез у прокариот
3.2 Синтез у эукариот
3.3 Транспорт липидов
3.3.1 Транспорт у прокариот
3.3.2 Транспорт липидов у эукариот
3.4 Изменение в липидном составе под действием окружающей среды
4 Заключение
5 Список литературы
1 Введение
Ранние этапы становления жизни являются неизученными проблемами биологии. На сегодня ни одна из многочисленных теорий не в состоянии дать устраивающего всех варианта происхождения жизни.
Идеальные условия для образования и сколь-нибудь длительного существования нужных для биогенеза молекул могли быть созданы только при наличии комфортной среды, которая отличалась бы от агрессивного окружения. По всей вероятности такие условия были созданы при появлении липидов, которые при взаимодействии с водой образовали примитивные липосомальные микросферы. По своей пространственной организации замкнутая форма липидной мембраны соответствует наименьшему значению энергии Гиббса, то есть термодинамически выгодна по сравнению с другими возможными формами расположения молекул. Кроме того, конформационная специфика бислойной фосфолипидной оболочки соответствует жидкокристаллическому состоянию, что предусматривает автономность по отношению к окружающей среде и одновременно селективную и регулируемую связь с этим внешним окружением.
Этот уникальный вариант не мог не закрепиться в ходе последующей биологической эволюции и не создать предпосылок для формирования гомеостаза, как одного из основополагающих принципов жизни.
Биогенез мембраны начинается с процессов синтеза составляющих ее компонентов – белков, липидов, углеводов. Затем эти компоненты должны быть доставлены к месту назначения и там образовать нужные структуры.
2 Образование белков
Две главные проблемы касающиеся сборки мембранных белков.
1. Все закодированные в ядре белки синтезируются общим пулом рибосом. В связи с этим возникает вопрос: как отдельные мембранные белки доставляются к месту назначения? Чем отличаются белки плазматической мембраны от белков внутренней митохондриальной мембраны или от белков мембран эндоплазматического ретикулума? Эту сложную проблему сортировки можно решить только при наличии определенных сигнальных последовательностей в каждом полипептиде, а также соответствующего аппарата узнавания.
2. Каков истинный механизм встраивания мембранных белков в мембрану и как при этом достигается правильная их ориентация относительно мембранного бислоя? Требуют ли механизмы встраивания и ориентации также наличия определенных сигнальных элементов и систем узнавания и если да, то каковы они? Какие свойства обеспечивают при встраивании мембранных белков формирование правильной третичной, а также червертичной структуры в случае мультисубъедииичных ансамблей?
В поиске ответов достигнуты большие успехи. Существует идентифицируемая часть полипептидной последовательности, которая служит участком узнавания, или «сигналом», направляющим отдельный полипептид к мембране, в которую он встраивается. Эти сигнальные участки часто расположены на N-конце новосинтезированного полипептида и отщепляются специфическими сигнальными пептидазами после встраивания его в нужную мембрану. Для обозначения N-концевого сигнала различными авторами использовались следующие термины: сигнальный пептид, сигнальная последовательность, транзитный пептид, лидирующий пептид, пре-последовательность.
Процессы трансляции и встраивания белков в мембрану можно разделить в эксперименте. Для сборки мембранных белков в большинстве случаев необходима энергия, отличающаяся по величине от той, которая требуется для их трансляции на рибосоме. Связавшийся с мембраной-мишенью полипептид должен, кроме того, находиться в конформации, в которой может осуществляться его перенос через мембрану или встраивание в нее. Во многих случаях перенос белков через мембраны происходит от N-конца к С-концу, при этом необходимо, чтобы белок был по крайней мере частично развернут или слабо свернут.
Доставка каждого белка к месту назначения обеспечивается иерархией сигналов, закодированных в каждом полипептиде. Например, большинство белков, предназначенных для эндоплазматического ретикулума или митохондрий, синтезируется в виде предшественников большей молекулярной массы (пре-белков); на N-конце у них имеется дополнительная последовательность, которая отщепляется особыми протеолитическими ферментами, имеющимися в этих орган ел л ах. Такие первичные сигналы весьма разнообразны и необходимы для того, чтобы полипептиды были узианы при транслокации специфическими рецепторами в этих органеллах. Связывание с митохондриями происходит сразу после завершения трансляции. Однако для большинства белков, направляемых в
эндоплазматический ретикулум в клетках млекопитающих, наблюдается иная картина. Как видно из рис.1 после связывания белков с соответствующей органеллой должна произойти дальнейшая сортировка. Для этого нужна дополнительная информация, которая также должна быть закодирована в каждой полипептидной последовательности и может рассматриваться как вторичные сигналы. В нескольких случаях их удалось идентифицировать как сигнальные последовательности, физически отделенные от первичных, хотя, возможно, так бывает не всегда.
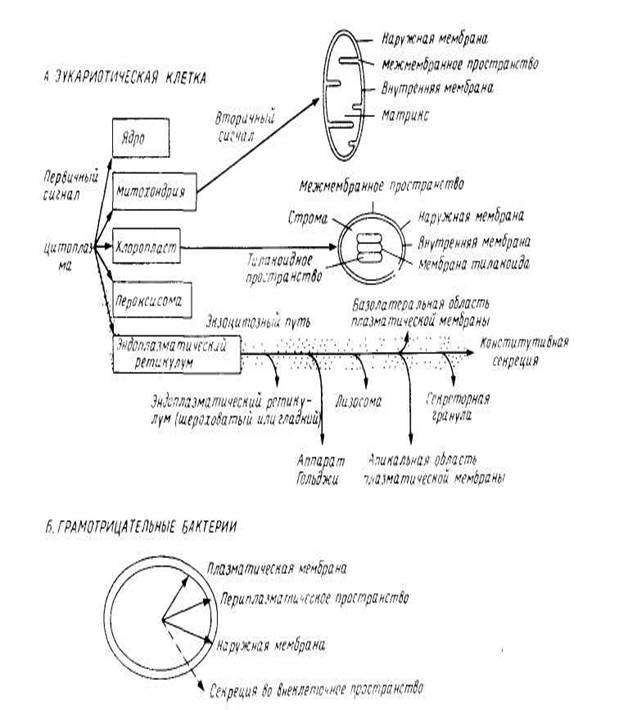
рис.1 Сортировка мембранных белков
биогенез мембрана белок липид
Особый интерес представляет процесс сборки мембранных белков, который целесообразно рассмотреть в связи с их сортировкой. На рис. 2 схематически показаны три общих механизма проникновения пептидного предшественника в мембрану. Механизмы А и Б являются вариантами схемы линейного вытеснения, согласно которой сигнальная последовательность направляет полипептид к переносящему устройству, которое включает в себя заполненный водой канал. Сигнальная последовательность может проходить прямо сквозь канал (механизм А) или оставаться связанной с мембраной, образуя, как показано на рис.2, петлю (механизм Б). В отсутствие какого-либо сигнала остановки процесса переноса полипептид будет транспортироваться через мембрану целиком. Однако, если внутри полипептида имеется второй сигнальный пептид, называемый стоп-сигналом переноса, то процесс останавливается и стоп-сигнал переноса становится трансмембранным сегментом зрелого мембранного белка. Фиксируя белок в мембране, стоп-сигнал переноса действует как сигнал сортировки. Если в белке имеются и другие сигналы начала и конца переноса, то будут образовываться следующие трансмембранные сегменты. Схема В на рис.2 иллюстрирует возможную роль самопроизвольного включения в мембрану гидрофобных элементов полипептидного предшественника. Этот механизм может реализовываться только тогда, когда включение в мембрану происходит после трансляции полипептида. Примером, подтверждающим существование этого механизма, является пробелок оболочки фага М13. Модель самопроизвольного включения может использоваться для объяснения механизма встраивания поперек мембраны амфифильных а-спиралей или β-структур. Этот процесс может также, конечно, быть белокзависимым.
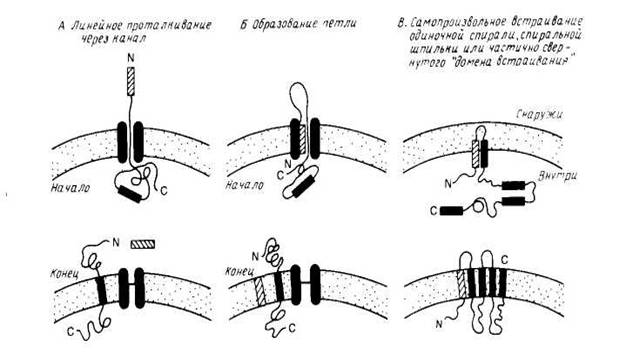
Рис2. Модели встраивания белка
Экспериментальные данные, которые однозначно свидетельствовали бы о существовании каналов, участвующих в сборке мембранных белков или в переносе белков через мембрану, отсутствуют. Известно, впрочем, что как на поверхности митохондрий, так и в эндоплазматическом ретикулуме имеются мембранные рецепторы, которые специфически узнают переносимые белки, и, возможно, именно они являются частью сложного аппарата, куда входит и канал, по которому перемещается белок.
Для того, чтобы перенос белков происходил со скоростью, близкой к скорости синтеза полипептида (1—10 остатков в 1 с), энергетический барьер не должен превышать примерно 18 ккал/моль. По полученным данным , две соседние спирали могут спонтанно встраиваться в бислой с образованием спиральной шпильки, и соответствующий выигрыш свободной энергии ~ 60 ккал/моль может стать движущей силой для частичного втягивания полярных и даже заряженных групп в липидный бислой. Однако, для переноса ионизированных и полярных групп из водного окружения в липидный бислой необходимо большее количество свободной энергии, н вряд ли модель спонтанного встраивания будет применима всегда, поскольку при сборке многих мембранных белков необходимо транспортировать через мембрану длинные, часто сильно заряженные полнпептидные цепи.
Тем не менее было показано, что некоторые небольшие мембранные белки включаются в лнпидные бислон спонтанно. К ним относятся цитохром bs с единственным гидрофобным якорем на С-конце и пробелок оболочки бактериофага М13, предположительно содержащий две трансмембранные спирали, которые, возможно, и встраиваются в бислой с образованием спиральной шпильки или петли. Пробелок оболочки содержит сигнальную последовательность из 23 остатков, обычно отщепляемую при сборке в цитоплазматической мембране Е. coli. Зрелый белок (50 аминокислотных остатков) имеет кислый N-конец, обращенный в периплазматическое пространство, трансмембранный сегмент и основный С-конец, обращенный в цитоплазму. Он спонтанно встраивается в фосфолипидные липосомы, причем скорость его сборки in vivo сильно замедляется, если в трансмембранном участке или на С-конце зрелого белка имеются мутации, что согласуется с моделью, в рамках которой два гидрофобных сегмента могут спонтанно встраиваться в липидный бислой в виде шпильки или петли. На рис. 3 представлена схема встраивания этого белка в мембрану. Интересно, что сборка пробелка оболочки вируса М13 может осуществляться и с помощью микросом млекопитающих, причем этот процесс требует АТР, возможно, для поддержания необходимой для транспорта конформации. Следует отметить, что этот белок не типичен для белков, сборка которых осуществляется на плазматической мембране Е. coli, поскольку он имеет отщепляемую сигнальную последовательность, и его сборка происходит независимо от функций генов secA, secY (prlA), необходимых для переноса белков внутрь плазматической мембраны или через нее.
Результаты исследования пробелка оболочки бактериофага М13 убедительно проиллюстрировали справедливость механизма самопроизвольного встраивания белков в мембрану без участия белков-посредников. Предполагается, что водорастворимый предшественник приобретает конформацию, обеспечивающую встраивание его в мембрану, при взаимодействии с бислоем. Эта обобщенная модель была предложена как часть «мембранной триггерной гипотезы». Необходимым условием нормального транспорта белка через мембрану является неполное его сворачивание в 3 и 4 структуры.
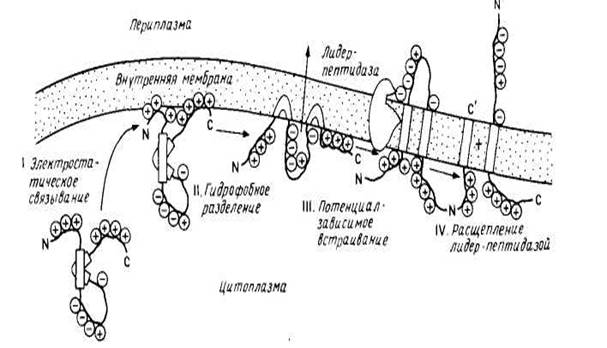
рис.3 модель самопроизвольного встраивания
2.1 Транспорт белков
Необходимо также рассмотреть механизмы с помощью которых белки доставляются к нужному месту. Есть несколько способов основным является экзоцитозный путь.
Экзоцитозным в эукариотических клетках называется путь, с помощью которого осуществляется транспорт белков, секретируемых клеткой или включаемых в наружную мембрану. Секретируемые белки синтезируются на связанных с мембранами рибосомах на цитоплазматической поверхности шероховатого эндоплазматического ретикулума и выводятся из клетки с помощью того же механизма, который используется для включения мембранных белков в эндоплазматический ретикулум.
Если водорастворимый белок не имеет вторичных сигналов сортировки, то он транспортируется к клеточной поверхности и секретируется с помощью «конститутивного» пути. Белки, транспортируемые этим путем, перемещаются из эндоплазматического ретикулума последовательно через различные компартменты комплекса Гольджи и в конце концов попадают на поверхность клетки. Они могут становиться компонентами цитоплазматической мембраны или, при наличии вторичных сигналов сортировки, оставаться в эндоплазматическом ретикулуме (рибофорин, цито-хром Р450) или в комплексе Гольджи (различные гликозилтрансферазы).
В комплексе Гольджи в ходе дальнейшей сортировки отделяются белки, секретируемые конститутивным путем (укороченный путь), от тех, которые направляются в лизосомы или концентрируются в секреторных гранулах, с помощью которых они затем секретируются при соответствующей стимуляции клетки (т. е. регулируемый секреторный путь). Кроме того, обнаружено, что в наружной мембране ядерной оболочки также могут синтезироваться мембранные гликопротеины, которые затем транспортируются с помощью экзоцитоза.Белки при транспортировке по экзоцитозному пути подвергаются посттрансляционным модификациям, в частности гликозилированию. Хорошо изучена компартментация процессинга N-связанного олигосахарида, что очень помогло определению различных компонентов комплекса Гольджи {цис-, медиальных и /пренс-цистерн).
Олигосахаридный предшественник с высоким содержанием маннозы присоединяется по местам гликозилирования в полипептиде (по остаткам аспарагина), когда белок находится внутри эндоплазматического ретикулума, а затем с помощью нескольких расположенных в различных компартментах ферментов осуществляется процесс созревания. Это позволяет следить за превращением белков, определяя состояние их гликозирования.
Исследования выявили несколько замечательных особенностей системы экзоцитозного транспорта.
1. Транспорт между различными компартментами органеллы осуществляется с помощью везикул, которые отпочковываются от «донорной» мембраны и потом сливаются с «акцепторной». Показано, что везикулы, участвующие в транспорте между компартментами комплекса Гольджи, являются «окаймленными», но соответствующий белок отличается от клатрина, окаймляющего эндоцитозные везикулы. Имеются веские данные в пользу того, что клатрин не является существенным компонентом экзоцитозного пути, хотя он, по-видимому, необходим для нормального роста некоторых штаммов дрожжей.
2. Для внутриклеточного транспорта необходим АТР, а также белковые компоненты цитозоля. Показано, что для транспорта между цистернами Гольджи, осуществляющегося при участии везикул, необходимо жирнокислотное производное ацил-СоА . Какую именно функцию выполняют указанные соединения в отпочковывании и слиянии везикул, неизвестно.
3. Роль олигосахарида как сигнала сортировки новосинтезированных гликопротеинов, по-видимому, непостоянна.
2.2 Сигнальные последовательности белков
У большинства белков, встроенных в мембрану эндоплазматического ретикулума или пересекающих ее, на N-конце имеется «короткоживущий» сигнальный пептид (от 15 до 30 аминокислотных остатков). Эта сигнальная последовательность непосредственно взаимодействует по крайней мере с двумя рецепторами, один из которых растворим (сигналраспознающая частица), а другой находится в мембране. Можно было бы ожидать, что аминокислотная последовательность этого сигнального пептида будет очень консервативной и примерно одинаковой у всех переносимых белков, но ожидания эти не оправдались. Эти сигнальные участки не отличаются постоянством ни в отношении длины, ни в отношении аминокислотной последовательности, а многочисленные опыты по мутагенезу показали, что они могут претерпевать значительные структурные изменения. Данные о том, что сигнальные пептиды содержат всю информацию, необходимую для транспорта белков через мембраны эндоплазматического ретикулума или внутрь их, были получены в опытах с химерными полипептидами. Присоединение N-концевой сигнальной последовательности к обычным цитоплазматическим белкам, например к глобину, приводило к тому, что они транспортировались в полость эндоплазматического ретикулума.
С точки зрения «сравнительной анатомии» N-концевых сигнальных последовательностей можно выделить три разных в структурном отношении участка: 1) положительно заряженный N-концевой участок (п-участок); 2) центральное гидрофобное ядро из 7—15 остатков (h-участок); З) С-концевой участок (с-участок), который является полярным и содержит сайт, узнаваемый сигнальной пепти-дазой, которая находится на стороне эндоплазматического ретикулума, обращенной в полость. Показано, что многочисленные случайные последовательности способны выполнять функцию нормального сигнального пептида у инвертазы дрожжей и детерминировать ее секрецию. Анализ этих случайных последовательностей показал, что решающим фактором является их гидрофобность. Приведены данные о гидрофобности и длине гидрофобных участков известных сигнальных пептидов эукариот и большинства гидрофобных участков, обнаруженных в цитозольных белках эукариот (многие из которых расположены на N-конце), а также известных трансмембранных якорных участков мембранных белков. Из этих данных видно, что h-область обладает свойствами, промежуточными между свойствами соответствующих участков цитозольных белков, с одной стороны, и типичных трансмембранных сегментов — с другой.
Очевидно, структурная специфичность для процесса узнавания не играет существенной роли. Однако необходимо помнить, что изменение свободной энергии менее чем на 5 ккал/моль (примерно такова энергия одной водородной связи) соответствует изменению сродства в 1000 раз. Такое различие в сродстве вполне может быть обусловлено тонкими различиями между функциональными и нефункциональными сигнальными последовательностями. Моделью рецептора сигнального пептида может служить растворимый фрагмент антигена гистосовместимости класса I, а именно HLA-A2, трехмерная структура которого известна. Этот белок связывается с пептидами — компонентами чужеродных антигенов, что является частью иммунного ответа. Область связывания пептида представляет собой большой желобок, открытый с одного конца и способный вмещать пептид из 20 аминокислотных остатков, если тот имеет форму а-спирали. О пептидах, которые могут связываться с HLA-A2, известно немного; показано, в частности, что близкородственный антиген гистосовместимости класса II проявляет высокое сродство к самым разным аминокислотным последовательностям. По-видимому, наиболее важными ббщими характеристиками пептидов, которые могут связываться с высоким сродством, являются вторичная структура и амфифильность. Стабилизации комплекса могут способствовать многочисленные взаимодействия в области связывания.
Известно, что относительно небольшие различия между сигнальными последовательностями порождают огромные различия в поведении белка. Например, если сигнальная последовательность не распознается сигнальной пептидазой, то белок чаще остается связанным с мембраной, чем секретируется, хотя есть и исключения из этого правила. Обычно сигнальные последовательности, которые служат также N-концевыми якорями, имеют более протяженный гидрофобный h-участок длиной около 20 аминокислотных остатков; этот участок необходим для остановки переноса и/или образования стабильного якоря в мембранном бислое . Примером такой сигнальной/якорной последовательности служит трансферриновый рецептор. Заметим, что в этом случае сигнальная последовательность расположена не иа N-конце, а на расстоянии более чем 50 аминокислотных остатков от него.
Известны также случаи, когда сигнальная последовательность закрепляет зрелый белок в противоположной ориентации, т. е. N-конец оказывается обращенным наружу. В качестве примера можно привести цитохром Р450 микросом крысы.
Каким-то образом эти сигналь-ные/якориые последовательности «проталкивают» свой N-коиец через мембрану и останавливают трансляцию, так что основная часть белка остается в цитоплазме. Отмечалось, что в некоторых из этих случаев сигнальные последовательности «старт/стоп» несут по крайней мере одни отрицательный заряд в п-области. Однако для встраивания указанных мембранных белков, как и белков обычного типа, используется одинаковый аппарат переноса — СРЧ. Возможно, наличие отрицательного заряда облегчает самопроизвольный или опосредованный белком перенос N-концевых остатков через мембрану.
Как мы уже отмечали, сигнальные последовательности не обязательно находятся на N-конце белковой молекулы и могут направлять перенос обоих фланкирующих домеиов, по крайней мере в случае искусственных гибридных белков. Уникальным примером такого рода является овальбумин, секреция которого детерминируется неотщепляемой внутренней сигнальной последовательностью. У многих мембранных белков эндоплазматического ретикулума неотщепляемые сигнальные последовательности тоже расположены в средней части полипептидиой цепи и играют роль трансмембраниых якорей. В качестве примера можно привести асиалогликопротеиновый рецептор. Внутренняя сигнальная последовательность этого белка использует тот же аппарат переноса, что и N-концевая последовательность; и действительно, в искусственных гибридах эта внутренняя сигнальная последовательность функционирует как обычная N-концевая последовательность. Примерами белков с внутренней неотщепляемой сигнальной последовательностью, которые имеют многочисленные трансмембраниые сегменты и N-конец которых находится на внутренней стороне мембраны, служат переносчик глюкозы и анионный переносчик белок полосы 3. Напротив, у опсина, тоже содержащего внутренний неотщепляемый сигнальный пептид, N-конец находится с наружной стороны мембраны. Этот внутренний сигнал (предположительно первый трансмембранный сегмент) протягивает гидрофильный аминокислотный домен (36 аминокислотных остатков) через мембрану, и, таким образом, его ориентация противоположна той, которая наблюдается в более общем случае при переносе полипептида, начиная с С-конца. Причина такого поведения опсина неизвестна; возможно, важную роль играет природа N-концевого пептида.
Итак, от небольших изменений в сигнальных последовательностях зависит, будет ли «белок-пассажир» секретироваться в полостьэндоплазматического ретикулума или он останется прикрепленным к мембране, и какой будет ориентация N-конца мембранного белка. Было показано, что существуют все возможные топологические варианты. Важным моментом является то, что во всех случаях сборка осуществляется при помощи одного и того же аппарата.
2.3 Стоп-сигналы переноса
Как отмечалось в предыдущем разделе, для неотщепляемых сигнальных последовательностей, которые играют роль N-концевых якорей в образовавшемся мембранном белке, характерно наличие относительно длинных гидрофобных участков. Отсюда следует, что перенос может останавливаться просто при наличии протяженного гидрофобного участка, который способен образовать трансмембранную а-спираль. В пользу такого предположения свидетельствуют некоторые экспериментальные данные. Например, с помощью рекомбинантной ДНК в среднюю часть белка £. coli, в норме секретирующегося через плазматическую мембрану, встраивали гидрофобные сегменты. Если их длина была не менее 16 аминокислотных остатков, то транспорт белка блокировался и он оставался присоединенным к плазматической мембране. Можно возразить, что в данном случае речь идет о бактериальной системе, но, как мы увидим ниже (см. раздел, посвященный сигнальным последовательностям бактерий), механизмы переноса в про- и эукариотических системах, по-видимому, сходны. Далее были сконструированы варианты G-белка вируса везикулярного стоматита с измененными мембранными доменами. Длина гидрофобного сегмента могла составлять не 20, а 8 остатков, при этом полипептид оставался трансмембранным, хотя транспорт в плазматическую мембрану блокировался. Таким образом, природа стоп-сигнала переноса точно не известна. Необходимо выяснить два вопроса: 1) участвуют ли в остановке процесса специфические белки аппарата переноса; 2) определяется ли остановка переноса гидрофобиостью стоп-сигнала или какими-то более тонкими факторами? Было показано, что участки стоп-сигнальиой последовательности, ответственные за блокирование переноса через эндоплазматический ретикулум, могут никак не влиять на транспорт через мембрану хлоропласта. Это означает, что упомянутые два процесса могут существенно различаться.
Определение старт- и стоп-сигналов подразумевает линейную схему переноса, начинающегося с N-конца; об этом свидетельствует поведение простых систем. Однако оказалось, что последовательности, которые блокируют перенос в одном случае, могут инициировать его в другом.
Следовательно важна не только последовательность но и ее окружение в полипептиде.
2.4 Сигнальные пептидазы
Для удаления временных N-концевых сигнальных пептидов необходимы специфические белки. Наиболее полно охарактеризованы сигнальные протеазы из Е. coli. Большинство экспортируемых белков Е. coli содержат сигнальный пептид, который отщепляется на периплазматической поверхности внутренней мембраны с помощью лидер-пептидазы. Для переноса белков через внутреннюю мембрану эта пептидаза не нужна, но она необходима для высвобождения экспортируемого белка из цитоплазматической мембраны. In vitro очищенный фермент мог функционировать, будучи включенным в липосомы. Специфичность расщепления весьма высока, но не определяется исключительно аминокислотной последовательностью вблизи сайта расщепления.
Сигнальная пептидаза, функционирующая в эндоплазматическом ретикулуме, имеет ту же специфичность, что и соответствующий фермент Е. coli, что неудивительно, если учесть сходство сигнальных последовательностей. Была очищена сигнальная пептидаза из микросом эукариот. Показано, что она ассоциирована с другими полипептидами, возможно имеющими отношение к механизму переноса.
У Е. coli имеется вторая сигнальная пептидаза, участвующая в процессинге пролипопротеинов. Эти полипептидные компоненты оболочки Е. coli замечательны тем, что при созревании их N-конец модифицируется с помощью глицерида. Пролипопротеи-новая сигнальная пептидаза также находится в цитоплазматической мембране. После отщепления сигнальный пептид остается в цитоплазматической мембране и разрушается с помощью мембра-носвязанного фермента протеазы IV.
В митохондриях и хлоропластах должно присутствовать несколько сигнальных пептидаз, поскольку процессинг происходит более чем в одном компартменте . Растворимую пептидазу из митохондриального матрикса удалось частично очистить, но охарактеризована она не полностью.
2.5.Белки необходимые для узнавания сигнальных последовательностей
1. Сигнал-распознающая частица (СРЧ). Это растворимый рибонуклеопротеиновый комплекс, состоящий из шести разных белков и молекулы. СРЧ необходима для инициации переноса. Она связывается с сигнальной последовательностью образующегося полипептида во время его синтеза на рибосоме. Для препролактина, например, константа диссоциации составляет 1 нМ [11%]. С помощью метода фотохимического сшивания был идентифицирован один из полнпептидов (54 кДа), непосредственно взаимодействующий с сигнальной последовательностью предшественника. По некоторым данным, полученным для бесклеточных систем, связывание СРЧ ингибирует трансляцию или вызывает ее задержку. Впрочем, не исключено, что этот феномен является артефактом. Во всяком случае, как было показано на модельных опытах, его не обязательно привлекать для объяснения кинетики переноса белков in vivo [11%]. Одна из вероятных функций СРЧ состоит в предотвращении неправильного свертывания образующегося полипептида, которое может блокировать перенос (например, из-за экранирования сигнальных последовательностей). Задержка трансляции должна уменьшать вероятность такого ошибочного свертывания и, следовательно, увеличивать эффективность переноса белков.
Некоторые небольшие белки (<8,5 кДа) транспортируются в эндоплазматический ретикулум независимо от СРЧ. В их число входят препропептид GLa лягушки, препромелиттин (оба они являются предшественниками секретируемых белков) и пробелок оболочки фага М13. Во всех этих примерах конформация предшественника такова, что белки должны оставаться способными к переносу даже в отсутствие СРЧ и рибосом.
2. Рецептор СРЧ, или стыковочный белок. Комплекс СРЧ/ рибосома/образующаяся полипептидная цепь транспортируется в шероховатый эндоплазматический ретикулум, преодолевая энергию сильного взаимодействия между СРЧ и мембраносвязанным рецептором СРЧ, называемым также стыковочным белком. Рецептор СРЧ содержит субъединицу с мол. массой 73 кДа, присоединенную N-концом к мембране. Вероятно, рибосома также связывается со специфическими рецепторами, присутствующими в мембране.
3. Рецептор сигнальной последовательности. Сигнальная последовательность на образующейся полипептидной цепи перемещается от СРЧ ко второму рецептору, находящемуся в мембране и называемому рецептором сигнальной последовательности. Об этом свидетельствуют результаты опытов по фотохимическому сшиванию, в которых используется метка, связанная с сигнальной последовательностью препролактина. Предполагаемый мембраносвязанный рецептор представляет собой гликопротеин с мол. массой 35 кДа. Возможно, он образует часть канала, через который осуществляется перенос. С помощью такого же подхода с использованием поперечной сшивки и синтетического сигнального пептида был обнаружен еще один кандидат на роль рецептора сигнальной последовательности (45 кДа). Связь между этими двумя белками неизвестна и функции их до конца не установлены. Как только образовавшаяся полипептидная цепь связывается с мем-браносвязанным рецептором, СРЧ и ее рецептор могут освободиться от рибосомы и принять участие в новом цикле. О предполагаемом канале, участвующем в переносе, ничего не известно; очистка его является довольно сложной задачей.
3 Липиды
После рассмотрения механизмов синтеза транспорта и встраивания мембранных белков необходимо рассмотреть и процессы происходящие с липидами мембран.
В мембранах эукариотических клеток обнаружено огромное количество разных липидов, причем они не распределены равномерно по разным клеточным мембранам. Эта неравномерность относится к распределению как полярных головок, так и ацильных хвостов. Например, степень ненасыщенности фосфолипидов в общем случае уменьшается при переходе от эндоплазматического ретикулума к комплексу Гольджи и затем к плазматической мембране. Для животной клетки среднее молярное отношение холестеролфосфолипиды равно 0,3— 0,4, при этом для плазматической мембраны оно гораздо выше (0,8—0,9), чем для других мембран, таких, как шероховатый эндоплазматический ретикулум (около 0,1). Кроме того, для многих мембран характерно неравномерное распределение липидов и по разным половинам бислоя.
Вопрос о механизме создания и поддержания такой асимметрии очень интересен и сложен. Напомним, что многие мембраны эукариотических клеток участвуют в эндоцитозе и экзоцитозе, при которых от одних мембран везикулы отпочковываются, а с другими сливаются. Этот процесс селективен для белков, которые транспортируются с помощью везикул, но почему-то не приводит к идентичности липидного состава вовлекаемых в него мембран. Причина этого явления неизвестна, выяснены лишь некоторые его аспекты. Рассмотрим, где синтезируются мембранные липиды, а затем обсудим, как они могут транспортироваться к месту назначения.
3.1 Прокариоты
У Е. coli весь синтез фосфолипидов протекает в плазматической мембране. В общих чертах этот процесс представлен на рис.4. Жирные кислоты синтезируются в виде предшественников, ковалеитно связанных с белком — переносчиком ацила, а затем включаются в мембрану путем ацилирования с образованием CDP-диацилглицерола. Два ацилирующих фермента рис.4 проявляют предпочтение к ненасыщенным жирнокислотным группам, находящимся в положении sn-2. На уровне CDP-диацилглицерола происходит разветвление процесса, после чего образуется фосфатидилсерин или фосфатидилглицерол рис.4. Механизм регуляции равновесного соотношения между фосфолипи-дами почти не изучен. В штаммах, в которых фермент, необходимый для биосинтеза кардиолипина, синтезируется в количествах, в 10 раз превышающих норму, наблюдается лишь незначительное увеличение содержания этого фосфолипида в мембране.
Аналогичные результаты были получены для фосфатидилсеринсинтазы и фосфатидилглицеролфосфатсинтазы . Сверхпродукция этих ферментов оказывает лишь небольшое влияние на фосфолипидный состав мембраны. С другой стороны, полное отсутствие фосфатидилглицеролфосфатсинтазы губительно для клетки, вероятно, потому, что кислые фосфолипиды необходимы для обеспечения предшественниками других биосинтетических реакций или участвуют в регуляции.
Получены мутанты, содержащие очень мало фосфатидилглицеролфосфатсинтазы, у которых единственным основным фосфолипидом является фосфатидилэтаноламин. Очищены некоторые ферменты биосинтетического пути, в том числе CDP-диглицеридсинтаза и фосфатидилсеринсинтаза. Первый фермент, как и ожидалось, связан с мембранами, но фосфатидилсеринсинтаза связана с рибосомами в бесклеточных экстрактах.
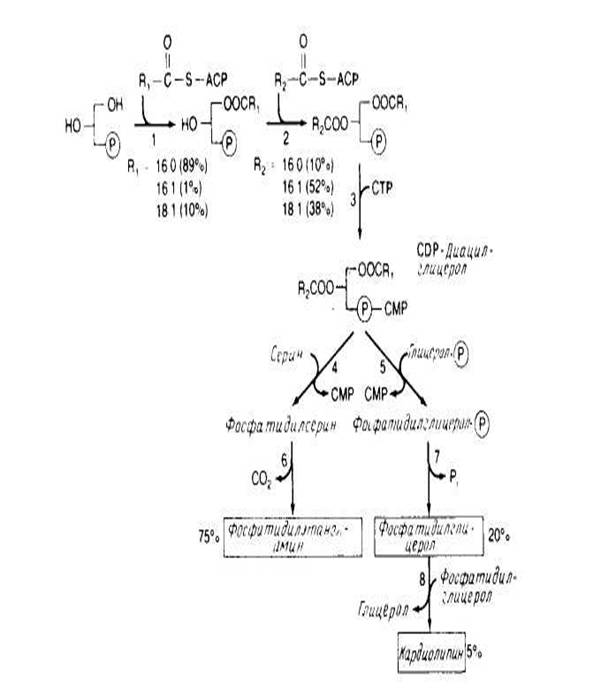
Рис.4 Синтез липидов у прокариот
3.2 У эукариот
На рис.5 представлена более сложная суммарная схема биосинтеза фосфолипидов в эукариотических клетках. Синтез жирных кислот происходит как в эндоплазматическом ретикулуме, так и в митохондриях. В качестве субстрата используется ацил-СоА, поскольку животные клетки не содержат эквивалента белку — переносчику ацила. Большинство ферментов, участвующих в биосинтезе фосфолипидов, локализовано на цитоплазматической стороне мембраны эндоплазматического ретикулума, но есть и исключения. В эукариотических клетках, как и в Е. coli, синтез фосфатидилглицерола и карднолипина может осуществляться через промежуточное образование CDP-диацилглицерола.
Однако ферменты, необходимые для этого, содержатся во внутренней мембране митохондрий, и эти фосфолипиды обнаруживаются только в митохондриях. Низшие эукариоты (например, дрожжи) тоже могут синтезировать фосфатидилсерин с помощью механизма, используемого Е. coli, но соответствующий фермент обнаружен как в эндоплазматическом ретикулуме, так и в митохондриях. В дрожжах на долю фосфатидилсерина приходится только около 5% всех фосфолипидов, но он является предшественником фосфатидилэтаноламина и фосфатидилхолина. Эукариоты, в том числе и дрожжи, могут синтезировать фосфа-тидилэтаноламин и фосфатидилхолин в ходе реакции с участием 1,2-днацилглицерола (рис. 5).
Это основной путь синтеза фосфолипидов в животных клетках. Затем специальные ферменты, находящиеся в эидоплазматическом ретикулуме высших эукариот (но ие дрожжей), катализируют реакцию обмена полярных головок, в результате чего образуется фосфатидилсерин. Эта реакция необходима высшим эукариотам для получения фосфатидилсерииа. После образования в эидоплазматическом ретикулуме фосфатидилсерин декарбоксилируется (по крайней мере в некоторых клетках) до фосфатидилэтаноламина. Фермент, катализирующий эту реакцию, фосфатидилсериндекарбоксилаза, находится в митохондриях. Следовательно, фосфатидилсерин может быть главным предшественником фосфатидилэтаноламина в клетке.
Для осуществления этой последовательности реакций должен происходить перенос липидов между эндоплазматическим ретикулумом и митохондрией. Фосфатидилэтаноламин может превращаться в фосфати-дилхолин с помощью одной или двух метилаз, содержащихся в эн-доплазматическом ретикулуме, но этот путь обычно не является главным.
Двумя липидными компонентами, которые локализуются в основном в плазматической мембране, являются сфингомиелин и холестерол. По-видимому, сфннгомиелин образуется в некоторых клетках путем переноса фосфатидилхолиновой группы от фосфати-дилхолина на церамид с помощью какого-то фермента, присутствующего в плазматической мембране. Напротив, несмотря на то, что холестерол концентрируется в основном в плазматической мембране, он синтезируется при участии ферментов, содержащихся в гладком эндоплазматическом ретикулуме и, возможно, в пероксисомах.
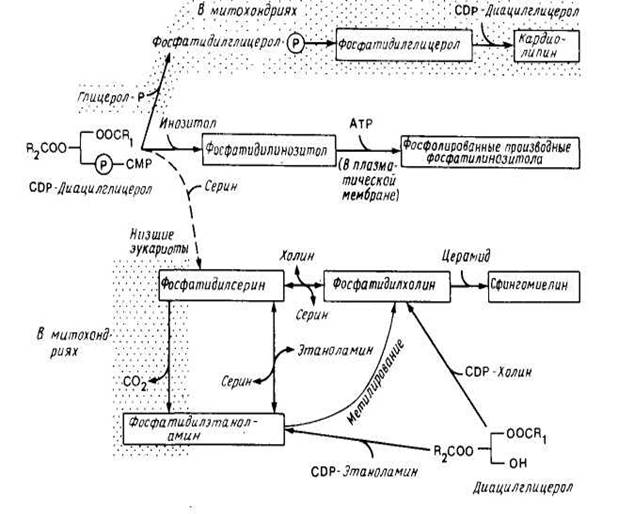
Рис.5. Синтез липидов у эукариот
3.3 Транспорт липидов
Перенос мембранных липидов от места их синтеза к месту назначения осуществляется при помощи двух процессов: 1) трансмембранного флип-флоп-перехода; 2) внутримембранного транспорта. Данные о скорости флип-флопа обсуждались в в связи с асимметрией мембраны. Скорость флип-флоп-перехода фосфолипидов особенно велика для тех мембран, в которых происходит биосинтез липидов; ее характерное время составляет величину порядка нескольких минут. Имеются данные о том, что этот процесс осуществляется при участии белков и, возможно, требует гидролиза АТР. Было также показано, что холестерол способен к быстрому спонтанному флип-флоп-переходу. Следовательно, транспорт через мембрану эндоплазматического ретикулума из цитозоля в просвет происходит довольно быстро.
В транспорте липидов от одной клеточной мембраны к другой участвуют несколько процессов. В разных случаях наиболее важным может оказаться какой-то один из них.
1. Самопроизвольный перенос липидов путем диффузии мономерных липидных единиц через водную фазу.
2. Диффузия липидов через постоянные или временные места соединения двух контактирующих мембран.
3. Транспорт с участием белков, катализируемый или белками, облегчающими высвобождение липидов из донорной мембраны, или липидсвязывающими белками.
4. Транспорт с участием везикул, при котором липиды, как и мембранные белки, транспортируются в ходе непрерывного отпочковывания и слияния с мембранами внутриклеточных везикул. Этот процесс может быть энергозависимым.
Рассмотрим вначале, что известно о самопроизвольной диффузии мембранных липидов между мембранами. Как показывают многочисленные исследования, липиды могут самопроизвольно перемещаться между моноламелляриыми везикулами или между фосфолипидными везикулами и биомембранамн. В большинстве случаев при этом происходит десорбция мономериых липидов с поверхности донорной мембраны и свободная диффузия через водную среду к акцепторной мембране. Лимитирующим этапом (по крайней мере при избытке акцепторных мембран) является высвобождение липидов из донорной мембраны. В этих условиях характерное время переноса зависит от величины свободной энергии десорбции. Ясно, что менее водорастворимые липиды (т. е. липиды с низкой критической концентрацией мицеллообразования) должны преодолевать при десорбции более высокий энергетический барьер, а следовательно, их перенос должен осуществляться медленнее. Скорость переноса зависит не только от гидрофобности переносимого липнда, но и от состава и физического состояния донорного бислоя.
Например, ганглиозид GMb находясь в фосфатидилхолиновых везикулах, существует в монодисперсном состоянии. Благодаря наличию гидрофильных полярных групп он не совершает флип-флоп-переходов через мембрану везикул, но характерное время его переноса везикулами составляет около 40 ч при 45 °С [153]. Напротив, нейтральные ганглиозиды, лишенные остатков сиаловой кислоты (например, асиало-GMi), образуют в везикулах гелеобразный кластер, и характерное время их переноса составляет около 500 ч. Смесь холестерола и фосфолипидов в везикулах тоже образует сложные фазы, и это может влиять на кинетику переноса холестерола. Стабилизация холестерола в мембране могла бы происходить за счет благоприятных взаимодействий со специфическими фосфо-липидами, например со сфингомиелином.
Перенос новосинтезированного холестерола из эндоплазматиче-ского ретикулума в плазматическую мембрану осуществляется всего за 10 мин. На процесс оказывают влияние агенты, блокирующие биоэнергетические реакции в клетке, например цианид. Эти и другие данные свидетельствуют о том, что внутриклеточный транспорт холестерола является энерогозависимым процессом и протекает при участии везикул. В принципе он может перевесить любой спонтанный перенос. Однако единого мнения на этот счет не выработано. Серьезной проблемой является то, что оценки доли холестерола, присутствующего в плазматической мембране, от общего его количества в клетке сильно варьируют (от 25 до 95%). На первый взгляд кажется, что количество холестерола, поступающего в некоторые клетки и выходящего из них, можно оценить, используя данные о скорости самопроизвольной диффузии мономеров, однако неясно, пригодны ли в данном случае механизм спонтанного переноса и указанные скорости.
Характерное время переноса фосфолипидов из фосфолипидных везикул гораздо больше, чем холестерола. Например, для дипальмитоилфосфатидилхолина оно составляет 83 ч при 37° в случае везикул из димиристоилфосфатидилхолина. Скорость переноса фосфолипидов с помощью этого механизма слишком мала, чтобы соответствовать реальным скоростям межмембранного транспорта.
3.3.1 Транспорт липидов у прокариот
Остановимся вначале на переносе фосфолипидов в случае грамотрицательных бактерий. Это относительно простая система, поскольку в ней имеются только две мембраны. Фосфолипиды синтезируются в плазматической мембране и должны транспортироваться в наружную мембрану. Имеются данные о том, что между этими мембранами осуществляется быстрый обмен липидами. Так, фосфатидилэтаноламин достигает наружной мембраны за характерное время - 3 мин. В этом процессе не участвуют белки, липиды или АТР, и каким-то образом зависит от протондвижущей силы. Липиды, внедрившиеся в наружную мембрану, могут также быстро перемещаться к внутренней мембране; это относится даже к фосфатидилхолину, который в норме не обнаруживается в Е. coli. Таким образом, процесс, по-видимому, не является специфическим. Тем не менее у мутантиого штамма с дефектной диацилглнцеролкиназой диацилглицерол накапливается в плазматической мембране и к наружной мембране не транспортируется.
Механизм липидного обмена между этими двумя мембранами неизвестен, но обычно полагают, что внутренняя и наружная мембраны имеют места соединения или области адгезии. По-видимому, именно через эти участки и происходит диффузия липидов. Здесь же могут концентрироваться белки, экспортируемые к наружной мембране.
3.3.2 Транспорт липидов у эукариот
Механизм распределения фосфолипидов в эукариотических клетках гораздо более сложен. Единственным фосфолипидом, который локализуется исключительно в месте своего синтеза, в митохондрии, является кардиолипин. Как показывают многочисленные эксперименты, внутриклеточный транспорт фосфолипидов осуществляется значительно быстрее, чем если бы он определялся самопроизвольной диффузией в водной среде.
Например, перенос новосинтезированного фосфолипида от эндоплазматического рети-кулума к митохондриям в клетках печени крысы происходит всего за несколько минут. Поскольку декарбоксилаза, катализирующая превращение фосфатидилсерина в фосфатидилэтаноламин, содержится в митохондриях, данные о скорости этой реакции могут использоваться для контроля за переносом фосфатидилсерина из эндоплазматического ретикулума в митохондрию. Показано, что скорость переноса весьма высока и процесс ингибируется азидом и агентами, блокирующими биоэнергетические реакции.
Обнаружено также, что новосинтезированный фосфолипид в течение нескольких минут транспортируется к плазматической мембране, хотя скорость переноса очень сильно зависит от самого липида и типа клетки. В некоторых случаях транспорт липидов блокируется ингибиторами биоэнергетических реакций (энергетическими ядами) и/или агентами, разрушающими цитоскелет. Общепринято, что скорость переноса липидов выше скорости диффузии.
Одним из способов, позволяющих облегчить перенос фосфолипидов между мембранами, является участие в этом процессе белков. Выделен целый ряд водорастворимых белков, участвующих в межмембранном переносе фосфолипидов in vitro. Все они имеют липидсвязывающие участки и образуют водорастворимые комплексы с фосфолипидами. Эти белки облегчают обмен липидов «один к одному» между мембранами путем переноса мономеров.
В одних случаях (например, для фосфатидилхолинспецифического белка переноса из печени быка) связывание липндов происходит с высокой специфичностью и сродством, в других сродство и специфичность относительно низки. В принципе при низком сродстве способность белка катализировать перенос липидов от одной мембраны к другой должна повышаться, поскольку белок при этом может возвращаться к донорной мембране, имея незанятый липидсвязывающнй участок. Белки, переносящие фосфолипиды, были выделены не только из животных клеток, но и из бактерий и клеток растений. Очищены белки, связывающиеся сгликолипидами и длинноцепочечными жирными кислотами,
К сожалению, функция белков, переносящих фосфолипиды, до сих пор не продемонстрирована in vivo, поэтому неясно, играют ли они физиологическую роль во внутриклеточном переносе фосфолнпидов. Не исключено, что этот перенос опосредуется везикулами, а возможно, функционируют оба механизма.
Важно понять, что так или иначе распределение фосфолипидов среди различных клеточных мембран детерминируется белками. Если распределение липидов равновесно, то оно определяется сродством белков, присутствующих в разных мембранах, к липидам. Смещение в распределении некоторых липидов из-за взаимодействия их с белками может влиять на распределение других липидов, например холестерола . Если распределение липидов неравновесно, то липидный состав разных мембран будет определяться скоростью транспортировки липидов к мембранам и от них, но и в этом случае кинетика процесса будет зависеть от участия в нем определенных белков.
3.4 Изменение состава липидов под действием условий внешней среды
Из всего сказанного ясно, что о механизме контроля липидного состава клеточных мембран известно очень мало. Одно из минимальных требований состоит в том, что мембранные липиды должны образовывать стабильный бислой, находящийся в жидкокристаллическом состоянии. Механизмы, обеспечивающие изменение липидного состава мембран в ответ на изменения условий окружающей среды, имеют многие организмы. Лучше всего изучен механизм тепловой адаптации Е. coli.
1. Тепловая адаптация Е. Coli.. При выращивании Е. coli в условиях низких температур наблюдается изменение жирнокислотного состава в сторону большего содержания ненасыщенных жирных кислот. Это способствует поддержанию мембраны в текучем состоянии и позволяет клеткам выжить при экстремальных температурах. Преобладающими остатками жирных кислот в Е. coli) являются пальмитоил (16:0), пальмитолеил (16:1) и цис-ваксеноил (18:1). При низких температурах в бислой включается больше дос-ваксеновой кислоты, а содержание пальмитолеиловой кислоты остается постоянным. Связано это с функционированием одного из двух ферментов, которые катализируют удлинение жнр-нокислотных цепей. При низких температурах фермент 3-кетоацил-АСР-синтаза II более активно превращает пальмитолеиловую кислоту в дос-ваксеновую, увеличивая пул ненасыщенных жнриых кислот, включаемых в фосфолипиды.
2. Адаптация к повышению давления. Жирнокнслотный состав барофильной морской бактерии NPT3 зависит от давления. При изменении давления в мембранах может также происходить фазовый переход, и эта адаптация, как и в предыдущем случае, позволяет организму выжить. При повышении давления содержание в мембранах ненасыщенных жирных кислот увеличивается, и благодаря особеиностям их упаковки мембрана остается в жидкокристаллическом состоянии. Аналогичные результаты были получены при изучении фосфолипидов митохондрий из клеток печени глубоководных океанических рыб.
4 Заключение
Клетки эукариот содержат много мембранных органелл и множество различных внутриклеточных мембран, каждая из которых обладает уникальным белковым и липидным составом. Любой мембранный белок, информация о синтезе которого заключена в ядре, должен безошибочно доставляться от места синтеза на риоо-соме, находящейся в цитоплазме, к месту назначения. Для этого используется сложная система сигнальных последовательностей, содержащихся в любой зрелой форме полипептида или предшественника, а также рецепторы внутри клетки, способные эти сигналы распознавать. Некоторые мембранные белки включаются в липидный бислой самопроизвольно, но в большинстве случаев правильная сборка белка внутри клеточной мембраны является энергозависимым процессом, который осуществляется с помощью специализированного аппарата. По-видимому, белки не могут включиться в клеточную мембрану до тех пор, пока они не приобретут частично развернутую конформацию. Разворачивание белков или поддержание их в развернутой конформацин, необходимой для переноса, возможно, осуществляются при участии АТР и специфических белков в цитоплазме.
В эукариотической клетке большинство липидов синтезируется в эндоплазматическом ретикулуме, и чтобы попасть к месту назначения, они должны пройти через соответствующие мембраны. Механизм транспорта липидов неизвестен, можно лишь предположить, что в нем участвуют везикулы и белки, связывающие липиды. Способ поддержания липидного состава различных внутриклеточных мембран тоже не установлен.
Литература
1.Р. Геннис «Биомембраны, молекулярная структура и функции» Москва «Мир» 1997 г.
2.В.К. Рубальченко «Структура и функции мембран» Киев 1988г.
3. Болдырев А.А. Лекции по биохимии мембран.
4. Финдлей Дж. Биохимия мембран. Методы. «Мир » 1990г.
Перепечатка материалов без ссылки на наш сайт запрещена