Курсовая работа: G-белки и их функция
Курсовая работа: G-белки и их функция
Содержание
Введение. 2
Из истории открытия С-белков. 8
Структура и свойства. 8
Связь с мембраной. 9
Стуктурно-функциональная организация G-белков. 9
Классификация по чувствительности к токеинам.. 10
Сопряжение с эффекторными системами. 10
Регуляция активности G-белков. 11
Аденилатциклаза. 12
Фосфолипазы.. 13
Протеинкиназы.. 14
Фосфодиэстеразы.. 16
Аденилатциклазная система. 17
Влияние бактериальных токсинов на активность аденилатциклазы (АДФ-рибозилирование G-белков) 20
Инозитолфосфатная система. 21
Участие белка кальмодулина в инозитолфосфатной передаче сигнала. 22
Саморегуляция системы.. 23
α-субъединица: общие свойства. 23
β и γ субъединицы: общая характеристика. 24
G-белки: βγ-субъединицы.. 25
ГТФ-связывающие белки образуют два основных семейства G-белков и низкомолекулярных ГТФ-связывающих белков. 28
Литература. 30
Введение
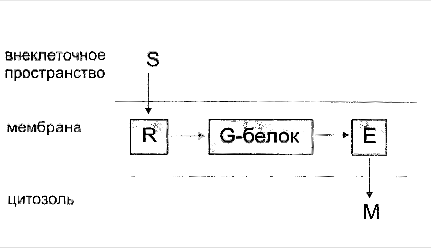
Сигнальные G-белки являются универсальными посредниками при передаче гормональных сигналов от рецепторов клеточной мембраны к эффекторным белкам, вызывающим конечный клеточный ответ. Когда семидоменная рецепторная молекула, локализованная в мембране сенсорной клетки, активируется какими-то изменениями во внешней среде, она претерпевает конформационные изменения. Последние детектируются
G-белками связанными с мембраной, которые, в свою очередь, активируют эффекторные молекулы в мембране. Часто это приводит к выделению вторичных мессенджеров в цитозоль.
Они являются объектом интенсивного изучения в связи с их участием во многих важных физиологических процессах. G-белки, участвующие в передаче сигнала, являются членами большого надсемейства гуанин-связывающих белков. G–белки - это прецизионные регуляторы, включающие или выключающие активность других молекул.
Примерно 80% первичных мессенджеров (гормоны, нейротрансмиттеры, нейромодуляторы) взаимодействуют со специфическими рецепторами, которые связаны с эффекторами через G-белки.
G-белки - белки, связывающие гуанозиновые нуклеотиды. G-белки, ассоциированные с рецепторами, связаны с мембраной. В неактивном состоянии они связаны с GDР. При связывании рецептора с лигандом ГДФ замещается на ГТФ, в результате чего происходит активация. Процесс этот сравнительно медленный, протекающий в течение секунд - десятков секунд.
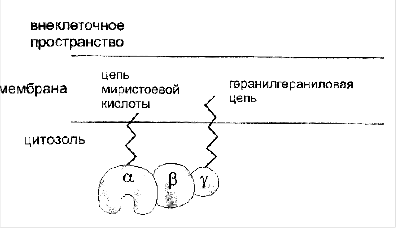
G-белки биологических мембран имеют гетеротримерную структуру. Они состоят из большой α-субъединиц (около 45 килодальтон - кДа), а также меньших β и γ-субъединиц, α-субъединица обладает ГТФ-азной активностью, в неактивной (выключенной) форме она связывает молекулу ГДФ на активном сайте. Субъединицы β и γ связаны между собой, и в физиологических условиях не могут быть диссоциированы. В неактивном состоянии βγ-комплекс непрочно связан с α-субъединицей. γ-субъединица связана с цитоплазматическим листком биологической мембраны геранил-гераниловой цепью (20 атомов углерода в цепи), близкой по структуре к холестерину. α-субъединица также связана с мембраной жирной кислотой с длиной цепи в 14 атомов углерода (миристоевая кислота). Такие связи обеспечивают то, что комплекс G-белка удерживается в плоскости мембраны, но в то же время способен легко двигаться в этой плоскости. Легко себе представить, как весь комплекс G-белка с присоединенным ГДФ перемещается в плоскости мембраны под действием тепловых сил, два семейства белков - гетеротримерные гуанозиннуклеотид связывающие белки (G-белки) и отдаленно родственные им гуанозинтрифосфатазы (ГТФ-азы) при связывании ГТФ могут включаться и активировать последующие компоненты передачи сигнала от поверхности клетки. Малые ГТФ-азы участвуют в контроле фундаментальных свойств клетки - полярности формы и процессов деления и дифференцировки. G-белки обычно регулируют более специализированные сигналы - продукцию вторичных мессенджеров. И те и другие способны гидролизовать GTР и таким образом выключать сигнал.
Поскольку β - и γ-субъединицы G-белков чрезвычайно консервативны, G-белки принято различать по их α-субъединицам. Кроме ГТФ-связывающего мотива, каждая последовательность Gальфа содержит как минимум один центр связывания дивалентных катионов, а также сайты ковалентной модификации бактериальными токсинами, катализирующими NAD-зависимые АДФ-рибозилтрансферазные реакции. G-белки, стимулирующие аденилатциклазу (Gs) или участвующие в фототрансдукции (Gt, трансдуцин) служат субстратами для АДФ-рибозилирования, катализируемого холерным токсином по одному из остатков аргинина, что приводит к блокированию деактивации этих белков. Gs, G-белок, ингибирующий аденилатциклазу, (Gi) и G-белок с пока еще неизвестной функцией (Go) АДФ-рибозилируются коклюшным токсином по остатку цистеина, расположенному у С-конца. Эта модификация препятствует взаимодействию между G-белком и рецепторами. Определена последовательность G-белка крысы (Gx), который оказался нечувствительным к коклюшному токсину.
G-белки - это регуляторные белки, связывающие при активации ГТФ. Лучше всего изучены G-белки, стимулирующие и ингибирующие аденилатциклазу (Gs - белки и Gi-белки соответственно). βı - адренорецепторы, β2 - адренорецепторы и D1 рецепторы сопряжены с белком Gs, и поэтому стимуляция этих рецепторов сопровождается активацией аденилатциклазы и повышением внутриклеточной концентрации цАМФ - классического второго (внутриклеточного) посредника. Конечный ответ в разных клетках различен и зависит от того, что представляет собой эффекторные фрагменты (фермент, ионный канал и пр) α2– адренорецепторы, М2-холинорецепторы и D2-рецепторы сопряжены с белком Gi, и стимуляция этих рецепторов приводит к снижению активности аденилатциклазы и внутриклеточной концентрации цАМФ. Изменения активности ферментов и других внутриклеточных белков и, соответственно, клеточных функций при этом противоположны тем, что наблюдаются при активации белка Gs. α1-адренорецепторы (как и М1-холинорецепторы), видимо, сопряжены с другим, пока еще мало изученным типом G-белка. Этот белок иногда обозначают Gq. Он активирует фосфолипазу С, катализирующую распад мембранных фосфолипидов, в частности - фосфатидилинозитол-4,5-дифосфата до ИЗФ и ДГА. Оба эти вещества являются вторичными посредниками.
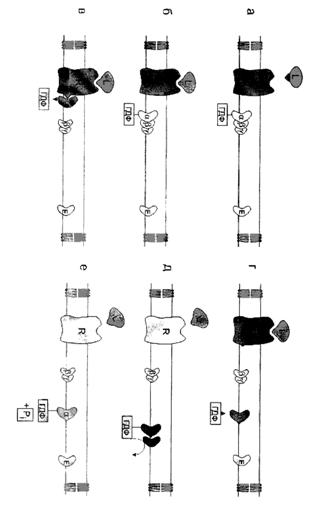
Связывание агониста (гормона, нейромедиатора и др.) с соответствующим рецептором приводит к белок-белковому взаимодействию между рецептором и G-белком и ускоряет диссоциацию ГДФ. В результате образуется короткоживущий комплекс агонист - рецептор - G-белок, не связанный ни с каким нуклеотидом. Связывание с этим комплексом молекулы ГТФ снижает сродство рецептора к G-белку, что приводит к диссоциации комплекса и высвобождению рецептора. Потенциально рецептор может активировать большое количество молекул G-белка, обеспечивая, таким образом, высокий коэффициент усиления внеклеточного сигнала на данном этапе. Активированная α-субъединица G-белка диссоциирует от βγ-субъединиц и вступает во взаимодействие с соответствующим эффектором, оказывая на него активирующее или ингибирующее воздействие.
α-субъeдиница с присоединенным с ней ГТФ способна взаимодействовать с эффектором в мембране - ферментами, такими, как аденилатциклаза, или, возможно, ионными каналами. Фермент может активироваться или ингибироваться, а ионный канал - открываться или закрываться. Конкретные примеры будут рассмотрены в последующих разделах. Взаимодействие с эффектором, однако, длится до тех пор, пока α - субъединица, являющаяся ГТФазой, удерживает ГТФ. Так что, очень вскоре присоединенный ГТФ гидролизуется до ГДФ. Когда это происходит, α - субъединица снова меняет свою конформацию и теряет способность активировать эффектор. После этого α-ГДФ взаимодействует с βγ-комплексом и снова образует тримерный комплекс, завершая, таким образом, цикл. Предполагают также, что комплекс из βγ-субъединиц тоже может (прямо или опосредованно) влиять на эффекторные ферменты.
Такими ферментами являются аденилатциклаза, фосфолипаза С. G-белки также регулируют работу К и Са²+-ионных каналов, К G-белкам относятся полипептид Gs, стимулирующий аденилатциклазу и регулирующий Са²+-ионные каналы, полипептид Gi, ингибирующий аденилатциклазу, и регулирующий К+-каналы в клетках тканей мозга, Gt, трансдуцин, участвующий в передаче светового сигнала, Golf специфичный белок обонятельных ресничек и др. Все G-белки являются гетеротримерами, состоящими из субъединиц α, β‚ и γ в порядке уменьшения молекулярной массы.
Впоследствии ГТФ, связанный с α-субъединицей G-белка, подвергается гидролизу, причем ферментом, катализирующим этот процесс, является сама α-субъединиц. Это приводит к диссоциации α-субъединицы от эффектора и реассоциации комплекса α-ГДФ с βγ - субъединицами. Спонтанная активация G-белка, связанного с ГДФ - весьма маловероятный процесс.
Этот же механизм лежит в основе гормональной регуляции фосфоинозитидспецифичной фосфолипазы С и фосфолипаза А2. Кроме того, было показано, что G-белки могут непосредственно активировать ионные каналы.
Лимитирующей стадией процесса восстановления исходного состояния G-белка является скорость диссоциации ГДФ от α-субъединицы G-белка. Скорость диссоциации увеличивается при взаимодействии G-белок-ГДФ с агонистсвязанным рецептором. Связывание ГТФ G-белком приводит, очевидно, к образованию комплекса агонист-рецептор-G-белок. Аналог GТР-СТР-γ-S и Мg2+ усиливает диссоциацию α-субъединицы из тримера G-белка. Однако следует заметить, что каталитическая субъединица аденилатциклазы из мембран мозга быка хроматографически соочищается с α - и β-субъединицами Gs-белка и вопрос диссоциации α-субъединиц из тримера G-белка для активации эффектора требует уточнения.
G-белки проявляют значительный полиморфизм. Каждая из форм субъединиц G-белка высокогомологична по структуре, близка по функциям, но отличается молекулярной массой и электрофоретической подвижностью. Особенно широк полиморфизм и наиболее изучен для αs и αi G-белков. Так из мозга человека выделено 11 форм ДНК, ответственных за синтез αs-субъединиц, четыре вида которых клонированы и, предполагается, что они определяют синтез четырех изоформ αs, в мозге человека. Для αi найдены, в основном, три изоформы αi1, αi2, αi3. Молекулярные массы изоформы αs находятся в пределах 42-55 кДа, а αi 39-41 кДа. Распределение молекулярных вариантов αi носит тканеспецифический характер: αi1 представлена, в основном, в мозге, αi2 обнаружена в нервной ткани и в клетках крови, αi3 представлена в периферических тканях и отсутствует в мозге. Распределение генов, кодирующих синтез трех изоформ αi по тканям примерно совпадает в ряду: человека, бык, крыса, мышь. Определение аминокислотной последовательности αi и αs показало, что изоформы αs или αi различаются в области С - и N - концевой последовательности, связывающихся с рецептором или эффектором. Предполагается, что полиморфизм α-субьединиц определяется многообразием рецепторов и их подтипов и разнообразием эффекторных систем.
αi-субъединицы Gi кодируются тремя различными структурными генами. Что касается изоформ α-субъединиц Gs-белков, то пока неясно, кодируются ли изоформы разными структурными генами или это продукт одного гена с последующим внутренним альтернативным сплайсингом исходного РНК-транскрипта, или множественность их результат посттрансляционной модификации. В настоящее время известно 9 структурных генов, кодирующих G-белки и 12 продуктов этих генов.
Из истории открытия С-белков
1. 1971г. - впервые показана необходимость ГТФ для стимуляции аденилатциклазы глюкагоном.
2. 1981г. - выделен белок Gt-трансдуцин, связывающий родопсин с фосфодиэстеразой с ГТФ фоторецепторов.
З. 1983г - выделен ГТФ-связываюший белок Gs, сопрягающий стимулирующие рецепторы с аденилатциклазой.
4. 1985-1988гт - показано, что фосфолипаза С и фосфолипаза А2 регулируются гормонами и нейротрансмиттерами через Gp-белки.
5. В настоящее время G-белки разделены на несколько типов: четыре Gs, три Gi, Go, Gz/x (центральная нервная система и селезенка), Gt (трансдуцин), Golf (обонятельные нейроэпителиальные клетки).
Структура и свойства
1. G-белки - гетеротримеры, в которых α-субъединица непрочно связана с димером β-γ.
2. Все известные α-субъединицы (мол. масса – 50кДа) гомологичны, и у большинства из них одинаковые (или очень сходные) b-субъединицы (мол. масса З5кДа) и γ-субъединицы (мол. масса 8кДа).
З. α-субъединица определяет специфичность связывания G-белка с рецептором и эффектором, уникальна для каждого G-белка.
4. α-субъединица связывает и гидролизует ГТФ (ГТФ-аза).
5. α-субъединица содержит высоко консервативный домен связывания и гидролиза ГТФ (18 аминокислот из 350-395).
6. Выявлены участки связывания гуаниновых нуклеотидов и участки взаимодействия с рецепторами (С-конец) и βγ-димерами (N-конец).
7. Выявлены участки АDР-рибозилирования (аргинин-202) при действии холерного токсина и коклюшного токсина.
Связь с мембраной
G-белки локализованы на внутренней поверхности плазматической мембраны. Первичная структура всех субъединиц G-белков не содержит гидрофобных, пронизывающих мембрану доменов.
1. Ассоциации G-белков с мембраной содействует ацилирование жирнокислотными радикалами. Выявлено два типа липидных модификаций субъединиц G-белков: миристоилирование и изопренилирование белковой цепи.
2. Показано для α-субъединиц Go - и Gi-белков посттрансляционное миристоилирование со стороны N-конца.
З. Для βγ-субъединиц также показаны посттрансляционные модификации (ацилирование).
4. Выявлены три последовательные посттрансляционные модификации, ответственные за связывание ras-белков с мембраной.
5. Очищенные α-субъединицы проявляют гидрофильные свойства (без βγ - комплекса не могут связываться с искусственными фосфолипидными пузырьками).
Стуктурно-функциональная организация G-белков
G-белки (ГТФ-связывающие белки) - универсальные посредники при передаче сигналов от рецепторов к ферментам клеточной мембраны, катализирующим образование вторичных посредников гормонального сигнала. G-белки - олигомеры, состоящие из α, β и γ-субъединиц. Состав димеров βγ незначительно различаются в разных тканях, но в пределах одной клетки все G-белки, как правило, имеют одинаковый комплект βγ-субъединиц. Поэтому G-белки принято различать по их α-субъединицам. Выявлено 16 генов, кодирующих различные α-субъединицы G-белков. Некоторые из генов имеют более одного белка, вследствие альтернативного сплайсинга РНК.
Каждая а-субъединица в составе G-белка имеет специфические центры:
связывания ГТФ или ГДФ;
взаимодействия с рецептором;
связывания с βγ-субъединицами;
фосфорилирования под действием протеинкиназы С;
взаимодействия с ферментом аденилатциклазой или фосфолипазой С.
В структуре G-белков отсутствуют α-спиральные, пронизывающие мембрану домены. G-белки относят к группе "заякоренных" белков.
Классификация по чувствительности к токеинам
1. ХТ (холерный токсин) приводит к постоянной активации аденилатциклазы (подавляя ГТФ-азную активность Аs-субъединицы)
2. КТ (коклюшный токсин) тоже вызывает АDР-рибозилирование α-субъединицы. Однако в этом случае модификация G-белка препятствует его взаимодействию с рецепторами, поэтому при активации рецептора А не ингибируется.
По чувствительности к холерному и коклюшному токсинам G-белки можно разбить на четыре группы: чувствительные только к холерному токсину (Gs), только к коклюшному (Gi и Go), субстраты обеих токсинов (Gt) и G-белки, α-субъединицы которых не чувствительны ни к одному из токсинов.
Сопряжение с эффекторными системами
ГТФ-связываюшие белки управляют несколькими мембранными ферментами и рядом ионных каналов.
Вероятно с G-белками взаимодействует цитоскелет, благодаря чему гормоны регулируют секрецию и эндоцитоз. Из мембранных и внутриклеточных мишеней G-белков лучше всего изучены аденилатциклаза и фосфодиэстераза ГМФ сетчатки глаза, активируемые, соответственно, Gzx и трансдуцином. Эти два фермента принципиально отличаются друг от друга по структуре и механизму их регуляции G-белками.
В отношении активации других G-белок зависимых систем ясности нет. G - белки опосредуют не только активирующее, но и ингибирующее действие агонистов на внутриклеточные эффекторные системы. G-белок зависимое ингибирование показано для аденилатциклазы, потенциалправляемых кальциевых каналов, фосфолипазы С, Nа/К-АТФазы.
Исходя из данных, можно предположить, что существует два механизма G-белок зависимого ингибирования аденилатциклазы. Один из них обусловлен действием βγ-субъединиц и, видимо, одинаков для всех G-белков, т.к βγ-субъединицы у них сходные. Второй механизм заключается в специфическом ингибировании аденилатциклазы α-субъединицей белка Gi.
Регуляция активности G-белков
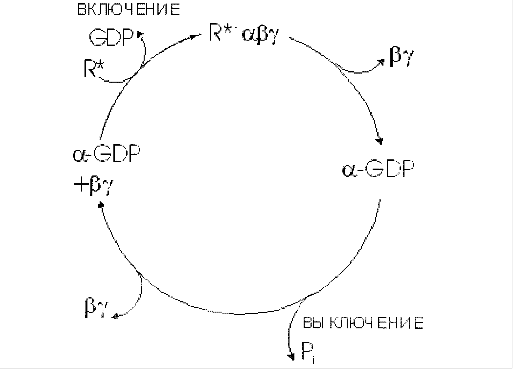
Различают неактивную форму G-белка - комплекс αβγ-ГДФ и активированную форму αβγ-ГТФ. Активация G-белка происходит при взаимодействии с комплексом активатор-рецептор, изменение конформации G-белка снижает сродство α-субъединицы к молекуле ГДФ и увеличивает к ГТФ.
Замена ГДФ на ГТФ в активном центре G-белка нарушает комплементарность между α-ГТФ и βγ-субъединицами. Рецептор, связанный с сигнальной молекулой, может активировать большое количество молекул G-белка, таким образом обеспечивая усиление внеклеточного сигнала на этом этапе.
Активированная α-субъединица G-белка (α-ГТФ) взаимодействует со специфическим белком клеточной мембраны и изменяет его активность. Такими белками могут быть ферменты аденилатциклаза, фосфолипаза С, фосфодиэстераза цГМФ, Nа+-каналы, K+-каналы.
Следующий этап цикла функционирования G-белка - дефосфорилирование ГТФ, связанного с α-субъединицей, причём фермент, катализирующий эту реакцию, - сама α-субъединица.
Дефосфорилирование приводит к образованию комплекса α-ГДФ, который не комплиментарен специфическому белку мембраны (например аденилатциклазе), но имеет высокое сродство к βγ-протомерам. G-белок возвращается к неактивной форме - αβγ-ГДФ. При последующей активации рецептора и замене молекулы ГДФ на ГТФ цикл повторяется снова. Таким образом, αβγ-субъединицы G-белков совершают челночное движение, перенося стимулирующий или ингибирующий сигнал от рецептора, который активирован первичным посредником (например, гормоном), на фермент, катализирующий образование вторичного посредника.
Некоторые формы протеинкиназ могут фосфорилировать α-субъединицы G - белков. Фосфорилированная α-субъединица не комплиментарна специфическому белку мембраны, например аденилатциклазе или фосфолипазе С, поэтому не может участвовать в передаче сигнала.
Аденилатциклаза
Фермент аденилатциклаза, катализирующий превращение АТФ в цАМФ - ключевой фермент аденилатциклазной системы передачи сигнала. Аденилатциклаза обнаружена во всех типах клеток.
Фермент относят к группе интегральных белков клеточной мембраны, он имеет 12 трансмембранных доменов. Внеклеточные фрагменты аденилатциклазы гликозилированы. Цитоплазматические домены аденилатциклазы имеют два каталитических центра, ответственных за образование цАМФ - вторичного посредника, участвующего в регуляции активности фермента протеинкиназы А.
На активность аденилатциклазы оказывают влияние как внеклеточные, так и внутриклеточные регуляторы. Внеклеточные регуляторы (гормоны, эйкозаноиды, биогенные амины) осуществляют регуляцию через специфические рецепторы, которые с помощью α-субъединиц G-белков передают сигналы на аденилатциклазу. αs-субъединица (стимулирующая) при взаимодействии с аденилатциклазой активирует фермент, αi-субъединица (ингибирующая) ингибирует фермент. В свою очередь, аденилатциклаза стимулирует проявление ГТФ-фосфатазной активности α-субъединиц. В результате дефосфорилирования ГТФ образуются субъединицы αs-ГДФ и αi-ГДФ, некомплементарные аденилатциклазе.
Из 8 изученных изоформ аденилатциклазы 4 - Са2+-зависимые (активируются Са2+). Регуляция аденилатциклазы внутриклеточным кальцием позволяет клетке интегрировать активность двух основных вторичных посредников цАМФ и Са2+.
Фосфолипазы
Фосфолипазы - ферменты класса гидролаз, катализирующие катаболизм глицерофосфолипидов. Различают фосфолипазы секреторные, входящие в состав панкреатического сока, и клеточные фосфолипазы. Клеточные фосфолипазы А1, А2, D, С различаются по специфичности к отщепляемой группе. Все фосфолипазы - кальций зависимые ферменты.
Фосфолипаза С - фермент, гидролизующий фосфоэфирную связь в глицерофосфолипидах. В клетках человека идентифицировано 10 изоформ фосфолилазы С, различающихся по молекулярной массе, локализации, способу регуляции, субстратной специфичности. В структуре всех изоформ фосфолипазы С отсутствуют гидрофобные домены, которые могли бы обеспечить их взаимодействие с мембраной. Однако некоторые формы фосфолипазы С связаны с мембраной с помощью гидрофобного "якоря" - ацильного остатка миристиновой кислоты или за счёт взаимодействия с поверхностью бислоя. Каталитическая активность всех изоформ фосфолипазы С зависит от ионов кальция.
Большинство фосфолипаз С специфично в отношении фосфатидилинозитолов и практически не гидролизует другие типы фосфолипидов. Активный фермент может гидролизовать до 50% от общего количества фосфатидилинозитолов клеточной мембраны. При гидролизе фосфатидилинозитол-4,5-бисфосфата (ФИФ2) образуются гидродугты диацилглицерол (ДАГ) и инозитол-1,4,5-трифосфат (ИФ3), служащие вторичными посредниками в трансмембранной передаче сигнала по инозитолфосфатному пути.
Протеинкиназы
Все полярные сигнальные молекулы, действующие на клетку-мишень через мембранные рецепторы, осуществляют свою биологическую функцию путём фосфорилирования специфических белков и ферментов, регулирующих метаболизм в клетке. Фосфорилирование изменяет (увеличивает или уменьшает) их активность. Катализируют фосфорилирование белков (протеинов) протеинкиназы по аминокислотным остаткам серина, треонина, тирозина. Протеинкиназы могут быть субъединицей мембранного рецептора, например тирозиновая протеинкиназа рецептора инсулина, активность которой регулируется гормоном. другая группа - протеинкиназы, регулируемые вторичными вестниками гормонального сигнала (цАМФ, цГМФ, Са, ДАГ), например протеинкиназа А, протеинкиназа С, протеинкиназа G, кальмодулинзависимые протеинкиназы и др.
1. Протеинкиназы А
Протеинкиназы А (цАМФ-стимулируемые) участвуют в аденилатциклазной системе передачи сигнала. Протеинкиназа А состоит из 4 субъединиц R2C2 - двух регуляторных субъединиц (R2) и двух каталитических (C2). Комплекс R2C2 не обладает ферментативной активностью.
Комплекс R2C2 разными способами прикрепляется к мембране. Некоторые формы протеинкиназы А "заякориваются" с помощью алифатического остатка миристиновой кислоты каталитических субъединиц. Во многих тканях протеинкиназа А связана с "заякоренным" белком АКАРs (от англ. сАМР - dependent protein kinase anchoring proteins). АКАРs имеет центр связывания для регуляторных субъединиц протеинкиназы А. С помощью белка АКАРs протеинкиназа А связывается с мембраной в области локализации ферментов, катализирующих образование цАМФ (аденилатциклаза) или его гидролиз (фосфодиэстераза), а также белков, в регуляции активности которых фермент принимает участие, например потенциалзависимые Са2+-каналы.
Регуляторные субъединицы протеинкиназы А имеют специфические центры для связывания цАМФ. Присоединение цАМФ к регуляторным субъединицам приводит к изменению конформации последних и снижению сродства каталитическим субъединицам С, происходит диссоциация по схеме:
цАМФ4 + R2C2 = цАМФ4R2 + С + С
Субъединицы С представляют собой активную форму протеинкиназы А, которая катализирует реакции фосфорилирования белков по серину и треонину. Каталитические субъединицы С у разных типов протеинкиназ А не идентичны, они различаются прежде всего специфичностью в отношении белков-субстратов.
2. Протеинкиназы С
Протеинкиназы С участвуют в инозитолфосфатной системе передачи сигнала. Фермент состоит из двух функционально различных доменов - регуляторного и каталитическоо. Регуляторный домен содержит 2 структуры ("цинковые пальцы"), образованные фрагментами пептидной цепи, богатыми цистеином, и содержащими два иона цинка. "Цинковые пальцы" участвуют в связывании: диацилглицерола. Другой фрагмент регуляторного домена имеет высокое сродство к Са2+. Повышение концентрации кальция в цитозоле увеличивает сродство протеинкиназы С к фосфатидилсерину мембраны. Транслокация протеинкиназы С к мембране позволяет ферменту связаться с ДАГ, который ещё больше повышает сродство протеинкиназы С к ионам кальция. Наиболее распространённые изоформы протеинкиназы С активируются Са2+, диацилглицеролом и фосфатидилсерином.
Каталитический домен имеет центр, связывающий АТФ и белок-субстрат. Активная форма фермента протеинкиназы С фосфорилирует белки по остаткам серина и треонина. Снижение концентрации ионов кальция в клетке нарушает связь протеинкиназы С с фосфатидилсерином и диацилглицеролом, фермент переходит в неактивную форму и отделяется от мембраны.
3. Протеинкиназы G
В отличие от протеинкиназы А, протеинкиназа G присутствует не во всех тканях, её обнаруживают в лёгких, мозжечке, гладких мышцах и тромбоцитах. Изоформы протеинкиназы G могут быть связаны с мембраной или находиться в цитоплазме. Растворимая протеинкиназа G состоит из двух идентичных субъединиц, каждая из которых имеет два центра для связьвания цГМФ. Присоединение цГМФ к регуляторным центрам вызывает конформационные изменения субъединиц и повышает каталитическую активность фермента. Протеинкиназа G, подобно протеинкиназе А и С, специфична в отношении определенных белковых субстратов, которые она фосфорилирует по остаткам серина и треонина.
Фосфодиэстеразы
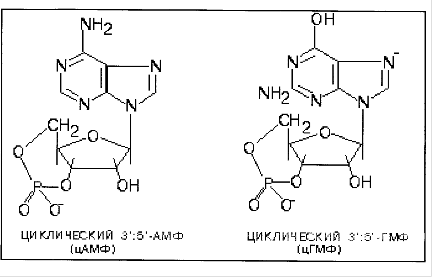
Фосфодиэстеразы - ферменты, катализирующие превращение цАМФ или цГМФ в неактивные метаболиты АМФ или ГМФ. Фосфодиэстеразы, снижая концентрации вторичных посредников, разрывают цепь превращений, вызванных активатором рецептора.
Фосфодиэстеразы присутствуют в клетках тканей в 2 формах: в форме растворимого белка и мембранносвязанного. Формы фермента, связанные с мембраной, в разных тканях составляют 5-40%. В одной и той же ткани могут присутствовать разные формы фосфодиэстеразы, различающиеся по сродству к субстратам, молекулярному весу, заряду, регуляторным свойствам и локализации в клетке.
Фосфодиэстеразы циклических нуклеотидов не обладают абсолютной специфичностью, поэтому, как правило, одна и та же форма фермента способна гидролизовагь как цАМФ, так и цГМф. Однако скорости гидролиза этих двух нуклеотидов под действием одной и той же фосфодиэстеразы могут значительно различаться. Это зависит от того, какая фосфодиэстераза присутствует в клетке - более специфичная в отношении цАМФ или более специфичная к цГМФ, от соотношения концентраций цАМФ и цГМФ в клетке и от действия регуляторов фосфодиэстеразы.
В большинстве тканей присутствует фосфодиэстераза - 1, более специфичная к цАМФ, активируемая Са2+, комплексом 4Са2+-кальмодулин и цГМФ.
Аденилатциклазная система
При участии аденилатциклазной системы реализуются эффекты сотни
различных по своей природе сигнальных молекул гормонов, нейромедиаторов, эйкозаноидов.
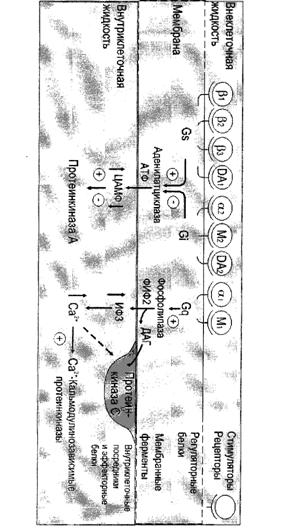
Функционирование системы трансмембранной передачи сигналов обеспечивают белки: Rs-рецептор сигнальной молекулы, которая активирует аденилатциклазу, и Ri-рецептор сигнальной молекулы, которая ингибирует аденилатциклазу; Gs-стимулирующий и Gi-ингибирующий аденилатциклазу белки; ферменты аденилатциклаза (АЦ) и протеинкиназа А (ПКА).
Последовательность событий, приводящих к активации аденилатциклазы:
связывание активатора аденилатциклазной системы, например гормона (Г) с рецептором Rs, приводит к изменению конформации рецептора и увеличению его сродства к Gs-белку. В результате образуется комплекс [Г] [R] [G-ГДФ] ;
присоединение [Г] [R] к G-ГДФ снижает сродство α-субъединицы Gs-белка к ГДФ и увеличивает сродство к ГТФ. ГДФ замещается на ГТФ;
это вызывает диссоциацию комплекса. Отделившаяся субъединица α, связанная с молекулой ГТФ, обладает сродством к аденилатциклазе:
[Г] [R] [G-ГТФ] = [Г] [R] + α-ГТФ + βγ
взаимодействие α-субъединицы с аденилатциклазой приводит к изменению конформации фермента и его активации, увеличивается скорость образования цАМФ из АТФ;
конформационные изменения в комплексе [α-ГГФ] [АЦ] стимулируют повышение ГТФ-фосфатазной активности α-субъединицы. Протекает реакция дефосфорилирования ГТФ, и один из продуктов реакции - неорганический фосфат (Р) отделяется от α-субъединицы, а комплекс [α-ГДФ] сохраняется; скорость гидролиза определяет проведения сигнала;
образование в активном центре α-субъединицы молекулы ГДФ снижает его сродство к аденилатциклазе, но увеличивает сродство к βγ-субъединицам. Gs-белок возвращается к неактивной форме;
если рецептор связан с активатором, например гормоном, цикл функционирования Gs-белка повторяется. Активация протеинкиназы А (ПКА)
молекулы цАМФ могут обратимо соединяться с регуляторными субъединицами ПКА
присоединение цАМФ к регуляторным субъединицам (R) вызывает диссоциацию комплекса С2R2 на комплекс цАМФ4R2 и С + С
активная протеинкиназа А фосфорилирует специфические белки по серину и треонину, в результате изменяются конформация и активность фосфорилированных белков, что приводит к изменению скорости и направления регулируемых ими процессов в клетке.
концентрация цАМФ в клетке может регулироваться, она зависит от соотношения активностей ферментов аденилатциклазы и фосфодиэстеразы.
Большую роль в регуляции внутриклеточной сигнальной системы играет белок АКАРs. "Заякоренный" белок АКАРs участвует в сборке ферментных комплексов, включающих не только протеинкиназу А, но и фосфодиэстеразу и фосфопротеинфосфатазу.
Каскадный механизм усиления и подавления сигнала. Передача сигнала от мембранного рецептора через G-белок на фермент аденилатциклазу служит примером каскадной системы усиления этого сигнала. Одна молекула, активирующая рецептор, может "включать" несколько G-белков и затем каждый активирует несколько молекул аденилатциклазы с образованием тысяч молекул цАМФ. На этом этапе сигнал усиливается в 10²-10³ раз. Образующийся цАМФ "включают" другой фермент - протеинкиназу А, усиливая сигнал ещё в 1000 раз. Фосфорилирование ферментов протеинкиназой. А ещё больше усиливает сигнал, в результате суммарное усиление равно 106-107 раз. Таким образом, по механизму каскадного усиления одна молекула регулятора способна изменить активность миллионов других молекул.
Но для любой из систем трансмембранной передачи сигнала клетка имеет другую систему, подавляющую этот сигнал. Каждый из этапов в ферментном каскаде находится под контролем специальных подавляющих этот сигнал механизмов. Например, длительное действие гормона приводит к десенсибилизации мембранных рецепторов: они либо инактивируются, либо вместе с гормоном погружаются в клетку посредством эндоцитоза. В результате десенсибилизации рецепторов степень активации аденилатциклазной системы снижается. Если в клетке длительное время повышена концентрация цАМФ (повышена активность протеинкиназы А), может происходить фосфорилирование кальциевых каналов, что приводит к повышению концентрации Са²+ в клетке. Калций активирует Са²+-зависимую фосфодиэстеразу, катализирующую превращение цАМФ в АМФ. В результате инактивации протеинкиназы А (R2С2) снижается скорость фосфорилирования специфических ферментов. Завершает "выключение" системы фосфопротеинфосфатаза, дефосфорилирующая фосфопротеины.
Влияние бактериальных токсинов на активность аденилатциклазы (АДФ-рибозилирование G-белков)
Для изучения функционирования G-белков аденилатциклазной системы были использованы экзогенные бактериальные яды холерный и коклюшный токсины. Токсины в экспериментальных условиях повышают активность аденилатциклазы практически во всех клетках организма; так, холерный токсин может стимулировать секрецию тиреоидных гормонов клетками щитовидной железы, стероидных гормонов клетками надпочечников, распад жиров в жировых клетках. Реакция разных клеток на холерный токсин вызвана повышением уровня цАМФ в этих клетках.
Холерный токсин - олигомерный белок. Одна из субъединиц фермент АДФ-рибозилтрансфераза; проникая в клетку, она катализирует присоединение АДФ-рибозы к αs-субъединице комплекса [αs - ГТФ] [АЦ] (этап активации аденилатциклазы).
NAD+ + [αs, - ГТФ] [АЦ] - [АДФ - рибозил - αs ГТФ] [АЦ] +никотинамид + Н+
АДФ-рибозилирование ингибирует проявление ГТФ-фосфатазной активности αs - субъединицы, не происходит дефосфорилированние ГТФ. Цикл функционирования G-белка останавливается на этапе активации фермента аденилатциклазы, отвечающего за образование цАМФ из АТФ. Фермент аденилатциклаза сохраняет повышенную активность в течение длительного времени.
Субъединица коклюшного токсина, проникая в клетку, катализирует АДФ-рибозилирование αi-субъединицы активированного Gi-белка(αi βγ-ГТФ).
NAD+ [αi βγ-ГТФ] - [АДФ-рибозил-αi βγ-ГТФ] + никотинамид + Н+
Модифицированная αi - субъединица сохраняет высокое сродство к βγ - субъединицам, те. Gi-белок теряет способность диссоциировать на αi - ГТФ и βγ-субъединицы. Таким образом, ингибирующий сигнал (αi-ГТФ) не достигает аденилатциклазы, значит в этом случае возможна только её активация при связывании с αs-ГТФ. Действие коклюшного токсина на клетки тканей всегда приводит к повышению уровня цАМФ.
Симптомы холеры и коклюша развиваются в результате действия токсинов, вырабатываемых соответствующими микроорганизмами.
Инозитолфосфатная система
Функционирование инозитолфосфатной системы трансмембранной передачи сигнала обеспечивают: R (рецептор), фосфолипаза С, Gрlс - белок, активирующий фосфолипазу С, белки и ферменты мембран и цитозоля.
Последовательность событий, приводящих к активации фосфолипазы С:
связывание сигнальной молекулы, например гормона с рецептором (R) вызывает изменение конформации и увеличение сродства к Gplc-белку.
образование комплекса [Г] [R] [Gрlс ГДФ] приводит к снижению сродства α-протомера G рlс белка к ГДФ и увеличению сродства к ГТФ. ГДФ заменяется на ГТФ.
это вызывает диссоциацию комплекса; отделившаяся α-субъединица, связанная с молекулой ГТФ, приобретает сродство к фосфолипазе С.
α-ГТФ взаимодействует с фосфолипазой С и активирует её. Под действием фосфолипазы С происходит гидролиз липида мембраны фосфатидилинозитол-4,5 - биофосфата (ФИФ2).
в ходе гидролиза образуется и выходит в цитозоль гидрофильное вещество инозитол-1,4,5-трифосфат (ИФ3). Другой продукт реакции диацилглицерол (ДАГ) остаётся в мембране и участвует в активации фермента протеинкиназы С (ПКС).
инозитол-1,4,5-трифосфат (ИФ3) связывается специфическими центрами Са2 - канала мембраны ЭР, это приводит к изменению конформации белка и открытию канала - Са²+ поступает в цитозоль. В отсутствие в цитозоле ИФ3 канал закрыт.
Активация протеинкиназы С.
• Повышение концентрации Са²+ в цитозоле клетки увеличивает скорость
взаимодействия Са²+ с неактивным цитозольным ферментом протеинкиназой С(ПКС) и белком кальмодулином, таким образом сигнал, принятый рецептором клетки, раздваивается.
• Связывание протеинкиназы С с ионами кальция позволяет ферменту вступать в кальций-опосредованное взаимодействие с молекулами "кислого" фосфолипида мембраны, фосфатидилсерина (ФС). Диацилглицерол, занимая специфические центры в протеинкиназе С, ещё более увеличивает её сродство к ионам кальция.
• На внутренней стороне мембраны образуется ферментативный комплекс - [ПКС] [Са²+] [ДАГ] [ФС] - активная протеинкиназа С, фосфорилирующая специфические ферменты по серину и треонину.
Участие белка кальмодулина в инозитолфосфатной передаче сигнала
В клетках многих тканей присутствует белок кальмодулин, который функционирует как внутриклеточный рецептор Са²+, он имеет 4 центра для связывания Са²+. Комплекс [кальмодулин] - [4Са²+] не обладает ферментативной активностью, но взаимодействие комплекса с различными белками и ферментами приводит к их активации.
Саморегуляция системы
Как и большинство систем трансмембранной передачи сигналов, инозитолфосфатная система имеет не только механизм усиления, но и механизм подавления сигнала. Присутствующие в цитозоле инозитол-1,4,5-трифосфат ((ИФ3) и диацилглицерол (ДАГ) в мембране могут в результате серии реакций опять превращаться в фосфатидилинозитол-4,5-бисфосфат (ФИФ2). Ферменты, катализирующие восстановление фосфолипида, активируются фосфорилированием протеинкиназой С.
Концентрация Са²+ в клетке снижается до исходного уровня при действии Са²+-АТФ-аз цитоплазматической мембраны и ЭР, а также Na+/Са²+-и Н+/Са²+-транслоказ (активный антипорт) клеточной и митохондриальной мембран.
Функционирование транслоказ Са²+ и Са²+-АТФ-аз может активироваться:
комплексом [камьмодулин] [4 Са] ;
протеинкиназой А (фосфорилированием);
протеинкиназой С (фосфорилированием). Понижение концентрации Са2 в клетке и диацилглицерола в мембране приводит к изменению конформации протеинкиiiазы С, снижению её сродства к фосфатидилсерину, фермеiтт диссоциирует в цитозоль (неактивная форма).
Фосфорилированные протеинкиназой С ферменты и белки под действием фосфопротеинфосфатазы переходят в дефосфорилированную форму.
α-субъединица: общие свойства
α-субъединица играет главную роль в функционировании G-белков. Она связывает ГТФ. Она обратимо взаимодействует с β и γ субъединицами, присоединяясь к ним, когда в центре находится ГДФ и диссоциируя, когда в центре ГТФ. При связывании ГТФ α субъединица активируется и приобретает способность регулировать эффекторные системы внутриклеточные. α субъединицы части G-белков могут подвергаться химическим модификациям. Под воздействием холерного и коклюшного токсинов происходит ФДФ-риболизирование белков по аргининовому и цистеиновому остатку на С-конце, в результате чего нарушается нормальное функционирование G-белков.
Кроме того, протеинкиназа С может фосфорилировать α-субъединицу очищенного G-белка, а in vivо белка Gz. По-видимому белки при этом инактивируются.
Большинство G-белков имеет α-субъединицы с молекулярным весом около 40 Кд.
β и γ субъединицы: общая характеристика
Бета и гамма субъединицы образуют комплекс друг с другом, распадающийся только в денатурирующих условиях. До конца их роль не ясна. В экспериментах с трансдуцином, а затем с белком Gi было показано, что субъединицы бета и гамма необходимы для взаимодействия G-белка с рецептором и замещения ГДФ на ГТФ.
Бета-гамма комплекс прочно связан с мембраной и служит якорем для α-субъединицы. При отделении α-субъединицы бета-гамма комплекс может переходить в цитоплазму.
Кроме связывания и ингибирования активности α-субъединицы бета - гамма комплекс в некоторых случаях оказывает прямое воздействие на эффекторные системы клетки. Он активирует фосфолипазу А2, взаимодействует с кальмодулином благодаря чему ингибирует активность аденилатциклазы мозга. G-бета-гамма комплекс ингибирует стимуляцию
АС1 по средством Gs-альфа.
АС2 стимулируется связыванием G-бета-гамма, но только в присутствии Gs - альфа.
АС3 также стимулируется G-бета-гамма. Калиевые каналы сердца открываются при связывании Gs-альфа и G-бета-гамма. Мембрана клеточная: стимуляция гидролиза фосфолипидов. Мембрана клеточная: изменение содержания цАМФ.
G-белки: βγ-субъединицы
Фосфорилирование рецепторов является одним из механизмов регуляции их активности. βγ-субъединицы G-белков могут осуществлять отрицательную обратную связь, активируя протеинкиназы, которые фосфорилируют рецепторы. Эти протеинкиназы называются GRК. К GRК протеинкиназам относятся родопсинкиназа и β-адренергическая киназа. Фосфорилирование приводит к удалению рецептора киназа. Например, мускариновые и адренорецепторы, фосфорилированные по серину и треонину на С-концевом домене, становятся мишенью для связывания арристина, что подготавливает их для удаления эндоцитозом. Обычно на С-конце рецептора есть несколько участков для фосфорилирования различными протеинкиназами. Известно, что слабый стимул (низкая концентрация агониста) активирует протеинкиназу А, а сильный стимул активирует b-АRК протеинкиназу, которая, фосфорилируя рецептор, прерывает передачу сигнала на аденилатциклазу и прекращает производство сАМР. Фосфорилирование, осуществляемое протеинкиназой А происходит тогда, когда занято 10% рецепторов. При этом фосфорилирование уже других, не занятых, рецепторов приводит к освобождению βγ-субъединиц и соответствующему фосфорилированию другой протеинкиназой b-АRК.
βγ-субъединицьт обеспечивают локализацию, эффективное связывание и дезактивацию α-субъединиц, регулируют сродство рецепторов к их активирующим лигандам, понижают способность GDР к диссоциации от субъединицы (стабилизация инактивированного состояния), открывает мускариновый К+-канал в сердце, закрывают Са2+ - канал в пресинаптической мембране, активируют фосфолипазу РLА2 и некоторые изоформы фосфолипазы С, регулируют сродство рецептора к агонисту.
ГТФ (GТР) - связывающие белки.
Передача сигналов от рецепторов, на внутриклеточные эффекторные системы осуществляется с помощью ГТФ-связывающих белков. ГТФ-связывающие белки активируются \ дезактивируются с помощью специального механизма - ГТФ-фазного цикла.
ГТФ-связывающие белки образуют два основных семейства G-белков и низкомолекулярных ГТФ-связывающих белков
Все эти белки сильно связывают ГТФ и превращают его в ГДФ, при этом происходит переход белка из активированного в неактивное состояние. Свойства ГТФ-связывающих белков. Основной структурной особенностью ГТФ-связывающих белков является наличие домена связывания гуаниновых нуклеотидов. Наличие соответствующих консенсусных аминокислотных последовательностей является однозначным указанием на принадлежность белка к данному семейству.
Литература
1. Биохимия.3-е издание (исправленное). М; Издательская группа ГЭОТАР-Медиа. 2006г.
2. Основы биохимии. А. Уайт, Ф Хендлер, Э. Смит, Р. Хилл, И. Леман.3 т. М; Мир1981 г.
3. Биохимия. Марри, Греннер. М; Высшая школа. 1993г.
4. Биохимия. Ленинджер, 1995г.
5. Биохимия. В.П. Комов., В.Н. Шведова. Дрофа; М: 2004г. .
6. Общая биохимия. Учебное пособие по биохимии. Курс лекций. М.Т. Генгин; 1997г.
7. Основы биохимии. Ю.Б. Филиппович. Издание второе, переработанное и дополненное. Высшая школа; М4, 1995г.
Перепечатка материалов без ссылки на наш сайт запрещена