Реферат: Аппарат Гольджи
Реферат: Аппарат Гольджи
СОДЕРЖАНИЕ
Введение. 3
Глава 1. Аппарат Гольджи: структура и функции. 5
1.1. Гольджи аппарат: структура. 5
1.2. Гольджи аппарат: функции. 8
Глава 2. Анализ деятельности аппарата Гольджи в клетке. 10
2.1. Анализ деятельности аппарата Гольджи в клетке. 10
2.2. Фибриллярные структуры.. 12
2.3. Аппарат Гольджи: сортировка белков и передача сигнала. 14
2.3. Аппарат Гольджи: молекулярный механизм функционирования. 15
Заключение. 18
Список литературы.. 21
Введение
Структуру, известную как комплекс Гольджи, впервые обнаружил в клетках животных в 1898 г. Камилло Гольджи, итальянский врач и цитолог. Детальное исследование данной структуры сделано позже с помощью электронного микроскопа.
Аппарат Гольджи содержится в цитоплазме почти всех эукариотических клеток, особенно в секреторных клетках животных. У дрожжей комплекс Гольджи выражен несколько хуже, обычно в виде особого отдела эндоплазматического ретикулума. Комплекс Гольджи представляет собой стопку уплощенных мембранных мешочков, так называемых цистерн, и связанную с ними систему пузырьков, называемых пузырьками Гольджи. На одном конце стопки мешочков постоянно образуются новые цистерны путем слияния пузырьков, отпочковывающихся, от гладкого эндоплазматического ретикулума. На другом конце стопки, на внутренней стороне завершается созревание цистерн и они вновь распадаются на пузырьки. Таким образом, цистерны в стопке постепенно перемещаются от наружной стороны к внутренней.
Функцией аппарата Гольджи является транспорт и химическая модификация поступающих в него веществ. Исходным субстратом для ферментов являются белки, поступающие в аппарат Гольджи из эндоплазматического ретикулума. После модификации и концентрирования, ферменты в пузырьках Гольджи переносятся к «месту назначения», например к месту образования новой почки. Наиболее активно этот перенос осуществляется с участием цитоплазматических микротрубочек.
Аппарат Гольджи является компонентом всех эукариотических клеток (практически единственное исключение - эритроциты млекопитающих). Он представляет собой важнейшую мембранную органеллу, управляющую процессами внутриклеточного транспорта . Основными функциями аппарата Гольджи являются модификация, накопление, сортировка и направление различных веществ в соответствующие внутриклеточные компартменты, а также за пределы клетки. Он состоит из набора окруженных мембраной уплощенных цистерн , напоминающих стопку тарелок. Каждая стопка Гольджи (у растений называемая диктиосомой ) обычно содержит от четырех до шести цистерн, имеющих, как правило, диаметр около 1мкм ( рис. 8-36 ). Число стопок Гольджи в клетке в значительной степени зависит от ее типа: некоторые клетки содержат одну большую стопку, тогда как в других имеются сотни очень маленьких стопок.
Со стопками Гольджи всегда ассоциирована масса мелких (диаметром приблизительно 60 нм) ограниченных мембраной пузырьков. Полагают, что эти пузырьки (пузырьки Гольджи ) переносят белки и липиды в аппарат Гольджи, транспортируют их из него и между остальными цистернами. Многие пузырьки являются окаймленными и покрыты клатрином или другим специфическим белком. Часто можно видеть, как такие окаймленные пузырьки отшнуровываются от цистерн Гольджи.
Аппарат Гольджи имеет две разные стороны: формирующуюся, или цис-сторону и зрелую, или транс-сторону Цис-сторона тесно связана с переходными элементами ЭР; транс-сторона расширяется, образуя трубчатый ретикулум, называемый транс-сетью Гольджи . Белки и липиды в составе небольших пузырьков попадают в стопку Гольджи с цис-стороны, а покидают ее, направляясь в различные компартменты, вместе с пузырьками, образующимися на транс-стороне. Переходя из одной стопки Гольджи в другую, эти молекулы претерпевают последовательные серии модификаций.
Глава 1. Аппарат Гольджи: структура и функции
1.1. Гольджи аппарат: структура
Описание структуры аппарата Гольджи тесно связано с описанием егоосновных биохимических функций, поскольку подразделение этогоклеточного компартмента на отделы производится преимущественно на основе локализации ферментов, расположенных в том или ином отделе.
Чаще всего в аппарате Гольджи выделяют четыре основных отдела: цис- Гольджи , медиал-Гольджи , транс-Гольджи и транс-Гольджи сеть ( TGN )
Кроме того к аппарату Гольджи иногда относят так называемыйпромежуточный компартмент, представляющий собой скопление мембранных пузырьков между эндоплазматическим ретикулумом и цис-Гольджи. Аппарат Гольджи является очень полиморфной органеллой; в клетках разных типов и даже на разных стадиях развития одной и той же клетки он может выглядеть по-разному. Основные его характеристики таковы:
1) наличие стопки из нескольких (обычно 3-8) уплощенных цистерн, более или менее плотно прилегающих друг к другу. Такая стопка всегда бывает окружена некоторым (иногда очень значительным) количеством мембранных пузырьков. В животных клетках чаще можно встретить одну стопку, в то время как в растительных клетках их обычно бывает несколько; каждую из них в таком случае называют диктиосомой. Отдельные диктиосомы могут быть связаны между собой системой вакуолей, образуя трехмерную сеть;
2) композиционная гетерогенность, выражающаяся в том, что постоянные (resident) ферменты неоднородно распределены по органелле;
3) полярность, то есть наличие цис-стороны, обращенной к эндоплазматическому ретикулуму и ядру, и транс-стороны,обращенной к поверхности клетки (это особенно характерно для секретирующих клеток);
4) ассоциация с микротрубочками и областью центриоли. Разрушение микротрубочек деполимеризующими агентами приводит к фрагментации аппарата Гольджи, однако его функции при этом существенно не затрагиваются. Аналогичная фрагментация наблюдается и в естественных условиях, во время митоза . После восстановления системы микротрубочек разбросанные по клетке элементы аппарата Гольджи собираются (по микротру-бочкам) в область центриоли,и реконструируется нормальный комплекс Гольджи.
Аппарат Гольджи (комплекс Гольджи) — мембранная структура эукариотической клетки, в основном предназначенная для выведения веществ, синтезированных в эндоплазматическом ретикулуме. Комплекс Гольджи был назван так в честь итальянского ученого Камилло Гольджи, впервые обнаружившего его в 1898 году.
Комплекс Гольджи представляет собой стопку дискообразных мембранных мешочков (цистерн), несколько расширенных ближе к краям и связанную с ними систему пузырьков Гольджи. В растительных клетках обнаруживается ряд отдельных стопок (диктиосомы), в животных клетках часто содержится одна большая или несколько соединенных трубками стопок.
В цистернах Аппарата Гольджи созревают белки предназначенные для секреции, трансмембранные белки плазматической мембраны, белки лизосом и т.д. Созревающие белки последовательно перемещаются по цистернам органеллы, в которых происходит их окончательное сворачивание, а также модификации — гликозилирование и фосфорилирование.
Аппарат Гольджи ассиметричен — цистерны располагающиеся ближе к ядру клетки (цис-Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки — везикулы, отпочковывающиеся от гранулярного эндоплазматического ретикулума (ЭР), на мембранах которого и происходит синтез белков рибосомами.
Разные цистерны Аппарата Гольджи содержат разные резидентные каталитические ферменты и, следовательно, с созревающими белками в них последовательно происходят разные процессы. Понятно, что такой ступенчатый процесс должен как-то контролироваться. Действительно, созревающие белки «маркируются» специальными полисахаридными остатками (преимущественно маннозными), по-видимому, играющими роль своебразного «знака качества».
Не до конца понятно, каким образом созревающие белки перемещаются по цистернам Аппарата Гольджи, в то время как резидентные белки остаются в большей или меньшей степени ассоциированы с одной цистерной. Существуют две взаимонеисключающие гипотезы, объясняющие этот механизм. Согласно первой (1), транспорт белков осуществляется при помощи таких же механизмов везикулярного транспорта, как и путь транспорта из ЭР, причем резидентные белки не включаются в отпочковывающуюся везикулу. Согласно второй (2), происходит непрерывное передвижение (созревание) самих цистерн, их сборка из пузырьков с одного конца и разборка с другого конца органеллы, а резидентные белки перемещаются ретроградно (в обратном направлении) при помощи везикулярного транспорта.
В конце концов от противоположного конца органеллы (транс-Гольджи) отпочковываются пузырьки, содержащие полностью зрелые белки.
В комплексе Гольджи происходит
1. О-гликозилирование, к белкам присоединяются сложные сахара через атом кислорода.
2. Фосфорилирование (присоединение к белкам остатка ортофосфорной кислоты).
3. Образование лизосом.
4. Образование клеточной стенки (у растений).
5. Участие в везикулярном транспорте (формирование трехбелкового потока):
6. созревание и транспорт белков плазматической мембраны;
7. созревание и транспорт секретов;
8. созревание и транспорт ферментов лизосом.
Аппарат Гольджи. Аппарат Гольджи (комплекс Гольджи) – это специализированная часть эндоплазматического ретикулума, состоящая из собранных в стопки плоских мембранных мешочков. Он участвует в секреции клеткой белков (в нем происходит упаковка секретируемых белков в гранулы) и поэтому особенно развит в клетках, выполняющих секреторную функцию. К важным функциям аппарата Гольджи относится также присоединение углеводных групп к белкам и использование этих белков для построения клеточной мембраны и мембраны лизосом. У некоторых водорослей в аппарате Гольджи осуществляется синтез волокон целлюлозы.
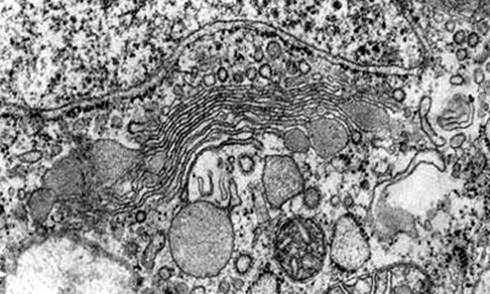
1.2. Гольджи аппарат: функции
Функцией аппарата Гольджи является транспорт и химическая модификация поступающих в него веществ. Исходным субстратом для ферментов являются белки, поступающие в аппарат Гольджи из эндоплазматического ретикулума. После модификации и концентрирования, ферменты в пузырьках Гольджи переносятся к «месту назначения», например к месту образования новой почки. Наиболее активно этот перенос осуществляется с участием цитоплазматических микротрубочек.
Функции аппарата Гольджи очень многообразны. К ним можно отнести:
1) сортировку, накопление и выведение секреторных продуктов;
2) завершение посттрансляционной модификации белков ( гликозилирование , сульфатирование и т.д.);
3) накопление молекул липидов и образование липопротеидов ;
4) образование лизосом ;
5) синтез полисахаридов для образования гликопротеидов, восков, камеди, слизей, веществ матрикса клеточных стенок растений
(гемицеллюлоза, пектины) и т.п.
6) формирование клеточной пластинки после деления ядра в растительных клетках;
7) участие в формировании акросомы ;
8) формирование сократимых вакуолей простейших.
Этот список, без сомнения, неполон, и дальнейшие исследования не только позволят лучше понять уже известные функции аппарата Гольджи, но и приведут к открытию новых. Пока самыми изученными с биохимической точки зрения остаются функции, связаные с транспортом и модификацией новосинтезированных белков.
Глава 2. Анализ деятельности аппарата Гольджи в клетке
2.1. Анализ деятельности аппарата Гольджи в клетке
Лизосомы – это маленькие, окруженные одинарной мембраной пузырьки. Они отпочковываются от аппарата Гольджи и, возможно, от эндоплазматического ретикулума. Лизосомы содержат разнообразные ферменты, которые расщепляют крупные молекулы, в частности белковые. Из-за своего разрушительного действия эти ферменты как бы «заперты» в лизосомах и высвобождаются только по мере надобности. Так, при внутриклеточном пищеварении ферменты выделяются из лизосом в пищеварительные вакуоли. Лизосомы бывают необходимы и для разрушения клеток; например, во время превращения головастика во взрослую лягушку высвобождение лизосомных ферментов обеспечивает разрушение клеток хвоста. В данном случае это нормально и полезно для организма, но иногда такое разрушение клеток носит патологический характер. Например, при вдыхании асбестовой пыли она может проникнуть в клетки легких, и тогда происходит разрыв лизосом, разрушение клеток и развивается легочное заболевание.
Митохондрии и хлоропласты. Митохондрии – относительно крупные мешковидные образования с довольно сложной структурой. Они состоят из матрикса, окруженного внутренней мембраной, межмембранного пространства и наружной мембраны. Внутренняя мембрана сложена в складки, называемые кристами. На кристах размещаются скопления белков. Многие из них – ферменты, катализирующие окисление продуктов распада углеводов; другие катализируют реакции синтеза и окисления жиров. Вспомогательные ферменты, участвующие в этих процессах, растворены в матриксе митохондрий.
В митохондриях протекает окисление органических веществ, сопряженное с синтезом аденозинтрифосфата (АТФ). Распад АТФ с образованием аденозиндифосфата (АДФ) сопровождается выделением энергии, которая расходуется на различные процессы жизнедеятельности, например на синтез белков и нуклеиновых кислот, транспорт веществ внутрь клетки и из нее, передачу нервных импульсов или мышечное сокращение. Митохондрии, таким образом, являются энергетическими станциями, перерабатывающими «топливо» – жиры и углеводы – в такую форму энергии, которая может быть использована клеткой, а следовательно, и организмом в целом.
Растительные клетки тоже содержат митохондрии, но основной источник энергии для yих клеток – свет. Световая энергия используется этими клетками для образования АТФ и синтеза углеводов из диоксида углерода и воды. Хлорофилл – пигмент, аккумулирующий световую энергию, – находится в хлоропластах. Хлоропласты, подобно митохондриям, имеют внутреннюю и наружную мембраны. Из выростов внутренней мембраны в процессе развития хлоропластов возникают т.н. тилакоидные мембраны; последние образуют уплощенные мешочки, собранные в стопки наподобие столбика монет; эти стопки, называемые гранами, содержат хлорофилл. Кроме хлорофилла, в хлоропластах имеются и все другие компоненты, необходимые для фотосинтеза.
Некоторые специализированные хлоропласты не осуществляют фотосинтез, а несут другие функции, например обеспечивают запасание крахмала или пигментов.
Относительная автономия. В некоторых отношениях митохондрии и хлоропласты ведут себя как автономные организмы. Например, так же, как и сами клетки, которые возникают только из клеток, митохондрии и хлоропласты образуются только из предсуществующих митохондрий и хлоропластов. Это было продемонстрировано в опытах на растительных клетках, у которых образование хлоропластов подавляли антибиотиком стрептомицином, и на клетках дрожжей, где образование митохондрий подавляли другими препаратами. После таких воздействий клетки уже никогда не восстанавливали отсутствующие органеллы. Причина в том, что митохондрии и хлоропласты содержат определенное количество собственного генетического материала (ДНК), который кодирует часть их структуры. Если эта ДНК утрачивается, что и происходит при подавлении образования органелл, то структура не может быть воссоздана. Оба типа органелл имеют свою собственную белок-синтезирующую систему (рибосомы и транспортные РНК), которая несколько отличается от основной белок-синтезирующей системы клетки; известно, например, что белок-синтезирующая система органелл может быть подавлена с помощью антибиотиков, тогда как на основную систему они не действуют.
ДНК органелл ответственна за основную часть внехромосомной, или цитоплазматической, наследственности. Внехромосомная наследственность не подчиняется менделевским законам, так как при делении клетки ДНК органелл передается дочерним клеткам иным путем, нежели хромосомы. Изучение мутаций, которые происходят в ДНК органелл и ДНК хромосом, показало, что ДНК органелл отвечает лишь за малую часть структуры органелл; большинство их белков закодированы в генах, расположенных в хромосомах.
Частичная генетическая автономия рассматриваемых органелл и особенности их белок-синтезирующих систем послужили основой для предположения, что митохондрии и хлоропласты произошли от симбиотических бактерий, которые поселились в клетках 1–2 млрд. лет назад. Современным примером такого симбиоза могут служить мелкие фотосинтезирующие водjросли, которые живут внутри клеток некоторых кораллов и моллюсков. Водоросли обеспечивают своих хозяев кислородом, а от них получают питательные вещества.
2.2. Фибриллярные структуры
Цитоплазма клетки представляет собой вязкую жидкость, поэтому можно ожидать, что из-за поверхностного натяжения клетка должна иметь сферическую форму, за исключением тех случаев, когда клетки плотно упакованы. Однако обычно этого не наблюдается. Многие простейшие имеют плотные покровы или оболочки, которые придают клетке определенную, несферическую форму. Тем не менее даже без оболочки клетки могут поддерживать несферическую форму из-за того, что цитоплазма структурируется с помощью многочисленных, довольно жестких, параллельно расположенных волокон. Последние образованы полыми микротрубочками, которые состоят из белковых единиц, организованных в виде спирали.
Некоторые простейшие образуют псевдоподии – длинные тонкие цитоплазматические выросты, которыми они захватывают пищу. Псевдоподии сохраняют свою форму благодаря жесткости микротрубочек. Если гидростатическое давление возрастает примерно до 100 атмосфер, микротрубочки распадаются и клетка приобретает форму капли. Когда же давление возвращается к норме, вновь идет сборка микротрубочек и клетка образует псевдоподии. Сходным образом на изменение давления реагируют и многие другие клетки, что подверждает участие микротрубочек в сохранении формы клетки. Сборка и распад микротрубочек, необходимые для того, чтобы клетка могла быстро менять форму, происходят и в отсутствие изменений давления.
Из микротрубочек формируются также фибриллярные структуры, служащие органами движения клетки. У некоторых клеток имеются бичевидные выросты, называемые жгутиками, или же реснички – их биение обеспечивает движение клетки в воде. Если клетка неподвижна, эти структуры гонят воду, частицы пищи и другие частицы к клетке или от клетки. Жгутики относительно крупные, и обычно клетка имеет только один, изредка несколько жгутиков. Реснички гораздо мельче и покрывают всю поверхность клетки. Хотя эти структуры свойственны главным образом простейшим, они могут присутствовать и у высокоорганизованных форм. В человеческом организме ресничками выстланы все дыхательные пути. Попадающие в них небольшие частички обычно улавливаются слизью на клеточной поверхности, и реснички продвигают их вместе со слизью наружу, защищая таким образом легкие. Мужские половые клетки большинства животных и некоторых низших растений движутся с помощью жгутика.
Существуют и другие типы клеточного движения. Один из них – амебоидное движение. Амеба, а также некоторые клетки многоклеточных организмов «перетекают» с места на место, т.е. движутся за счет тока содержимого клетки. Постоянный ток вещества существует и внутри растительных клеток, однако он не влечет за собой передвижения клетки в целом. Наиболее изученный тип клеточного движения – сокращение мышечных клеток; оно осуществляется путем скольжения фибрилл (белковых нитей) относительно друг друга, что приводит к укорочению клетки.
2.3. Аппарат Гольджи: сортировка белков и передача сигнала
Комплекс Гольджи функционирует на пересечении секреторных путей, осуществляя прием вновь синтезированных белков и липидов из ЭР , их ковалентную модификацию, а затем - сортировку продуктов реакций согласно их назначениям ( Рис. 1 gg ). В дополнение к этому, Гольджи возвращает некоторые компоненты в ЭР. Таким образом, Гольджи функционирует как в качестве области процессинга для синтезированных в ЭР гликопротеинов и гликолипидов, так и в качестве фильтрующей системы, отделяя белки, предназначенные для включения в плазматическую мембрану, от таковых, возвращаемых в ЭР [см. обзор Mellman и Simons, 1992 ]. Для исполнения указанных функций Гольджи организован в виде уплощенных цистерн со встроенными трансмембранными ферментами процессинга. Селективный транспорт-удержание различных видов белков и липидов в пределах данной системы опосредован цитозольными белками оболочки, аггрегирующими на поверхности мембран Гольджи, собирающими транспортируемый материал и способствующими изменению кривизны поверхности липидного бислоя таким образом, чтобы осуществить эндоцитоз и транспорт интермедиатов.
Ввиду того, что перемещающиеся внутрь или наружу транспортные интермедиаты часто преодолевают значительные расстояния, Гольджи неразрывно связан с цитоскелетом. В клетках млекопитающих положение Гольджи устанавливается микротрубочками.
Обилие актинсвязывающих белков в комплексе Гольджи, окружающих его подобно строительным лесам, также может способствовать пространственному контролю мембранного транспорта.
Каким образом Гольджи регулирует сортировку белков и эндоцитоз, а также координирует эти функции с цитоскелетом для достижения пространственного и временного контроля секреторного транспорта?
Два недавних исследования, дают пищу для рассуждений в данной области, предполагая, что трансмембранные транспортные белки и Cdc42 , организующая актин ГТФаза семейства Rho , конкурируют за один и тот же участок COPI , регулирующего секреторный транспорт.
2.3. Аппарат Гольджи: молекулярный механизм функционирования
Гептамерный цитозольный белковый комплекс, называемый COPI (мембранный комплекс Гольджи, коатомер) , в соединении с GTP-связывающим белком ARF 1 образует оболочку таким образом, что, будучи ассоциированным в мембраны Гольджи, предположительно содействует мембранному экзоцитозу и реакциям расщепления, связанным с мембранным транспортом Гольджи. Включение COPI в мембраны Гольджи требует присутствия ARF1, который работает по GTPазному циклу . ARF1-GTP осуществляет включение COPI в мембраны Гольджи, тогда как гидролиз GTP предположительно запускает высвобождение COPI из мембраны в цитозоль, что делает возможным включение COPI в периодические циклы сборки-разборки оболочки. Таким образом, ARF1 функционирует в качестве двойного переключателя, осуществляющего управление интеграцией COPI в мембраны и, следовательно, регулировку его функции.
Первоначально предполагалось, что связывающиеся с мембраной ARF1 и коатомер участвуют неселективно в формировании транспортных пузырьков. Данная модель предполагала наличие значительного потока транспортируемых веществ через секреторные пути и постулировала, что полимеризация коатомера, управляемая посредством циклизации ГТФ с помощью ARF1, обеспечивает механико-химическую энергию для образования пузырьков. В результате проведенных с тех пор разнообразных исследований указанная точка зрения была скорректирована. Активация ARF1 оказывает значительное влияние на фосфолипидный состав мембраны и стимулирует встройку , актина и других белков цитозоля в мембраны Гольджи. Это предполагает способность ARF1 облегчать процессы сортировки, эндоцитоза и стыковки мембран комплекса Гольджи.
Для фрагмента коатомера была также обнаружена способность связывать два остатка лизина в С-концевом мотиве трансмембранных белков , обеспечивающих циклический транспорт между Гольджи и ЭР и функционирующих, как предполагают, в качестве возвращающих в ЭР последовательностей [см. обзор Gaynor ea, 1998 ]. Взаимодействуя подобным образом с цитоплазматическими фрагментами транспортирующих белков, COPI может собирать транспортируемые вещества в везикулы и опосредовать сортировку транспортирных белков.
Что касается последней из указанных функций, предметом значительных дискуссий был вопрос о том, упаковывает ли мембраносвязанный COPI транспортируемые вещества в везикулы экзо- или эндоцитозного или обоих типов.
У дрожжей мутантные субъединицы COPI были идентифицированы по схеме, разработанной для обнаружения мутантов, неспособных удерживать/реутилизировать маркированные двумя остатками лизина молекулы при сохранении остальной части цикла.
В результате возникло предположение о том, что ассоциированный с содержащими дилизиновые мотивы трансмембранными транспортирующими белками COPI опосредует обратный транспорт. Однако, дальнейший анализ отдельных аллелей sec21 (гамма-COP) показал наличие зависящих от типа транспортируемого вещества селективных дефектов и при прямом транспорте [ Gaynor и др., 1998 ]. Более того, коатомер распознает также последовательности, родственные дилизиновым и диаргининовым, в цитоплазаматических фрагментах белков p24 , большого семейства потенциальных переносчиков, которыми изобилует Гольджи и для которых было показано участие в двунаправленном транспорте. С учетом этих, как и предыдущих биохимических и морфологических данных, подтверждающих роль коатомера в прямом транспорте, становится неясным направление (т. е., прямое или обратное) переноса везикул. Дополнительная возможность заключается в том, что опосредованная ARF1 ассоциация COPI с мембраной может служить для латерального разделения белков и липидов в отдельные группы, транспортируемые в дальнейшем прямым или обратным способом. Наличие данной функции было предложено в результате наблюдения того, что блокирование ассоциации COPI с мембраной у мутантов с ингибированием ARF1 либо посредством обработки брефельдином А (BFA) , предотвращающим активацию ARF1, само по себе не препятствует мембранному транспорту, но дестабилизирует его, что приводит к неселективности возврата белков в ЭР.
Заключение
Клетка, элементарная единица живого. Клетка отграничена от других клеток или от внешней среды специальной мембраной и имеет ядро или его эквивалент, в котором сосредоточена основная часть химической информации, контролирующей наследственность. Изучением строения клетки занимается цитология, функционированием – физиология. Наука, изучающая состоящие из клеток ткани, называется гистологией.
Существуют одноклеточные организмы, тело которых целиком состоит из одной клетки. К этой группе относятся бактерии и протисты (простейшие животные и одноклеточные водоросли). Иногда их также называют бесклеточными, но термин одноклеточные употребляется чаще. Настоящие многоклеточные животные (Metazoa) и растения (Metaphyta) содержат множество клеток.
Абсолютное большинство тканей состоит из клеток, однако имеются и некоторые исключения. Тело слизевиков (миксомицетов), например, состоит из однородной, не разделенной на клетки субстанции с многочисленными ядрами. Сходным образом организованы и некоторые животные ткани, в частности сердечная мышца. Вегетативное тело (таллом) грибов образовано микроскопическими нитями – гифами, нередко сегментированными; каждая такая нить может считаться эквивалентом клетки, хотя и нетипичной формы.
Некоторые не участвующие в метаболизме структуры тела, в частности раковины, жемчужины или минеральная основа костей, образованы не клетками, а продуктами их секреции. Другие, например древесина, кора, рога, волосы и наружный слой кожи, – не секреторного происхождения, а образованы из мертвых клеток.
Мелкие организмы, такие, как коловратки, состоят всего из нескольких сотен клеток. Для сравнения: в человеческом организме насчитывается ок. 1014 клеток, в нем каждую секунду погибают и замещаются новыми 3 млн. эритроцитов, и это всего одна десятимиллионная часть от общего количества клеток тела.
Обычно размеры растительных и животных клеток колеблются в пределах от 5 до 20 мкм в поперечнике. Типичная бактериальная клетка значительно меньше – ок. 2 мкм, а наименьшая из известных – 0,2 мкм.
Некоторые свободноживущие клетки, например такие простейшие, как фораминиферы, могут достигать нескольких сантиметров; они всегда имеют много ядер. Клетки тонких растительных волокон достигают в длину одного метра, а отростки нервных клеток достигают у крупных животных нескольких метров. При такой длине объем этих клеток небольшой, а поверхность очень велика.
Самые крупные клетки – это неоплодотворенные яйца птиц, заполненные желтком. Наибольшее яйцо (и, следовательно, наибольшая клетка) принадлежало вымершей громадной птице – эпиорнису (Aepyornis). Предположительно его желток весил ок. 3,5 кг. Самое крупное яйцо у ныне живущих видов принадлежит страусу, его желток весит ок. 0,5 кг.
Как правило, клетки крупных животных и растений лишь немногим больше клеток мелких организмов. Слон больше мыши не потому, что его клетки крупнее, а в основном потому, что самих клеток значительно больше. Существуют группы животных, например коловратки и нематоды, у которых количество клеток в организме остается постоянным. Таким образом, хотя крупные виды нематод имеют большее количество клеток, чем мелкие, основное различие в размерах обусловлено в этом случае все же большими размерами клеток.
В пределах данного типа клеток их размеры обычно зависят от плоидности, т.е. от числа наборов хромосом, присутствующих в ядре. Тетраплоидные клетки (с четырьмя наборами хромосом) в 2 раза больше по объему, чем диплоидные клетки (с двойным набором хромосом). Плоидность растения можно увеличить путем введения в него растительного препарата колхицина. Поскольку подвергнутые такому воздействию растения имеют более крупные клетки, они и сами крупнее. Однако это явление можно наблюдать только на полиплоидах недавнего происхождения. У эволюционно древних полиплоидных растений размеры клеток подвержены «обратной регуляции» в сторону нормальных величин несмотря на увеличение числа хромосом.
Мембраны Гольджи взаимодействуют также со множеством двигательных белков и белков, включая, спектрин , а также анкирин , облегчающий пространственное управление Гольджи мембранным транспортом, а также, вероятно, способный координировать сигнальные механизмы.
Каким образом могла бы быть организована и регулируема такая база? В течение нескольких лет было известно, что мембранный скелет , чьи компоненты включают актинсвязывающие белки спектрин и анкирин, ассоциирован с цитоплазматической поверхностью Гольджи. Структура этих "лесов" в совокупности с другими ассоциированными с Гольджи периферическими белками (включая многие из вышеупомянутых сигнальных молекул) сильно нарушается при обработке BFA . Это позволяет предположить, что их ассоциация с Гольджи либо непосредственно зависит от ARF1-ГТФ, либо находится в зависимости от комплексов, чья сборка инициируется активностью ARF-1. Недавно было показано, что активность ARF1 вызывает возрастание уровня содержания PIP2 в Гольджи посредством встраивания PI4K бета в мембраны Гольджи Поскольку генерирование PIP2 приводит к сборке актина и спектрина на мембране Гольджи, другие белки могут ассоциироваться и стабилизироваться в этой области, вероятно, ввиду возрастания эффективности сообщения между другими сигнальными молекулами и микротрубочками.
Таким образом, произведенный с помощью ARF1 PIP2 мог бы исполнять множество сходных сигнальных функций посредством встраивания молекул в мембрану, модулирования активности регуляторных молекул ARF, а также в качестве кофактора PLD.
Усилия исследователей в области мембранного транспорта, цитоскелета и преобразования сигнала необходимо объединить для детальной разработки роли регуляторных молекул и Cdc42 и COPI, в процессах сортировки и сигнального механизма комплекса Гольджи.
Список литературы
1. Албертс Б., Брей Д., Льюс Дж., Рэфф М., Робертс К., Уотсон Дж. Молекулярная биология клетки, т. 1. М., 1994
2. Власова З.А. Биология. Справочник студента – М., 2001
3. Воронцов Н. Н., Сухорукова Л. Н. Эволюция органического мира – М., 2002
4. Грин Н. Биология – М., 2003
5. Де Робертис Э. Новинский В., Саэс Ф. Биология клетки. М., Мир, 2001
6. Зегнбуш П. Молекулярная и клеточная биология. М., Мир, т2004
7. Камлюк Л.В. Биология в вопросах и ответах – Минск, 1994
8. Краткий справочник по химии, под ред. О. Д. Куриленко, 4 изд.. К., 1974
9. Лемеза Н.А. Пособие по биологии – Минск, 1998
10. Мамонтов С.Г. Биология – М., 2004
11. Меншуткин Н. А., Очерк развития химических воззрений, СПБ, 1888
12. Некрасов Б. В. Основы общей химии. М., 2001
13. Неницеску К. Д. Общая химия. Пер. с рум./ Под ред. Аблова А. В. – М.: Мир, 1968.
14. Свенсон К., Уэбстер П. Клетка. М., Мир, 2000.
15. Сидоров Е.П. Общая биология – М., 2003
16. Соловьев Ю. И., Эволюция основных теоретических проблем химии, М., 1971
17. Справочник химика, под ред. Б. П. Никольского, 2 изд., т. 1-6, М. - Л., 1965
18. Химическая энциклопедия: в 5 т. /Глав. ред. Кнунянц И.Л., Зефиров Н.С. – М.: Советская энциклопедия, Большая Российская энциклопедия, 1988–1998.
19. Хэм А., Кормак Д. Гистология, т. 1. М., 1982
20. Ченцов Ю.С., Поляков В.Ю. Ультраструктура клеточного ядра. М., Наука, 2004
21. Ярыгин В.Н. Биология – М., 2001
Перепечатка материалов без ссылки на наш сайт запрещена