Реферат: Фізіологія рослинної клітини
Реферат: Фізіологія рослинної клітини
МІНІСТЕРСТВО АГРАРНОЇ ПОЛІТИКИ УКРАЇНИ
ДНІПРОПЕТРОВСЬКИЙ ДЕРЖВАНИЙ АГРАРНИЙ УНІВЕРСИТЕТ
РЕФЕРАТ
НА ТЕМУ :
«ФІЗІОЛОГІЯ РОСЛИННОЇ КЛІТИНИ»
Виконав студент групи А- 3 -08
Гусак С.О.
Перевірила Професор Бессонова В.П.
Дніпропетровськ 2009
Зміст
1. Історія вивчення клітини і клітинна теорія
2. Сучасні методи вивчення клітини
3. Еволюція клітини
4. Ультра структура (мікроскопічна будова)
5. Біологічні мембрани їх будови і функції
6. Цитоскелет, мікротрубочки і мікрофіломенти,будова і функції
7. Ядро, його будова і функції
8. Ендоплазматична сітка, будова і функції
9. Рибосоми, будова і функції.
10. Мітохондрії, будова і функції
11. Пластиди, будова і функції
12. Мікросоми
13. Вакуолярна система
14. Апарат Гольджі, будова і функції
15. Будова і функції клітинних стінок
Література
1. Історія вивчення клітини і клітинна теорія
Людське око дозволяє бачити предмети розміром не менше 0.1 мм., що багато разів більше діаметра більшості клітин. Тому відкриття клітини і розвиток клітинної теорії тісно пов’язані з розвитком оптики і створення мікроскопа.
Перший мікроскоп, що складався з двох чи більше лінз, було винайдено голландськими механіками братами Янсен у 1600 році. Згодом його вдосконалив англійський фізик Роберт Гук. Виступаючи в Лондоні на зборах Королівського товариства він продемонстрував можливості свого приладу, популярність мікроскопа зросла.
У 1672 році італійський біолог і лікар Марчелло Мальпігі у своїй книзі « Анатомія рослин» докладно описує мікроскопічні структури рослинних тканин. В цей же час нідерландський натураліст Антоні Ван Левенгук, використовуючи мікроскоп, спостерігав і змалював сперматозоїди, еритроцити, найпростіших та інше.
За десятки років, що пройшли після відкриття Р.Гука, величезне число дослідників використовувало мікроскопи, накопичилась велика кількість повідомлень, описів і малюнків різних типів клітин, тканин і мікроорганізмів. Однак усі вченні того часу головною частиною клітин вважали їх стінки, не приділяючи належної уваги внутрішньоклітинним структурам. Так продовжувалося аж до ХІХ століття.
Так, у 1835 р. чеський натураліст Ян Пуркіньє вперше описує ядро тваринної клітини, вводить термін «протоплазма» і стверджує, що саме вона, а не клітина стінка, є живою речовиною. В 1826 р. російський вчений Карл Бер відкриває яйцеклітину і вивчає ембріогенез, що затвердило його засновника ембріології як науки. В 1830 р. англійський ботанік Роберт Браун описує ядро рослинної клітини, вказуючи на те що ця структура є обов’язковим компонентом усіх клітин. У 1846р. німецький ботанік Гуго фон Моль дає класифікацію тканинам рослин, та розподіляє поняття «протоплазма» і «клітинний сік».
Після визначення клітинної теорії вона почала інтенсивно розвиватися й уточнюватися. Наприкінці ХІХ століття німецький медик і біолог Рудольф Вірхов довів, що клітини утворюються з інших клітин шляхом їх поділу. За короткий час (1883-1898рр.) відкрито пластиди рослинних клітин, більш глибоко вивчено клітинний поділ і описані хромосоми, виявлені мітохондрії й апарат Гольджі.
У 30-40 року ХХ сторіччя з винаходом електронного мікроскопа почалася нова ера в розвитку біології. Стало можливим вивчити ті деталі клітин, про які раніше людина не мала уявлення. З цього часу в цитології і фізіології рослин наступив різкий перелом - замість роздільного вивчення структури клітин і їхніх функцій вони стали розглядатися в органічній єдності, що дозволило розкрити найважливіші фізіологічні процеси, які в них протікають.
2. Сучасні методи вивчення клітини
Нині існує безліч методів вивчення рослинних клітин. Найбільш широко використовуваний – класичний мікроскопічний метод, що полягає у вивченні живих і фіксованих клітин і тканин на тимчасових і постійних препаратах за допомогою звичайних світових мікроскопів. Широко використовуваним методом в електронній мікроскопії є метод «заморожування-сколювання». При цьому заморожену тканину (-196 0С) розколюють лезом і площина відколу в багатьох випадках проходить через гідрофобну зону мембран, що дозволяє по-новому глянути на мікроструктуру клітин і установити положення окремих макромолекул.
Для вивчення хімічного складу і функцій окремих клітинних структур з успіхом застосовується методи фракціонування клітин чи їхнього вмісту. Екстракти зруйнованих клітин ділять на функції, піддаючи їх високошвидкісному центрифугуванню. Крупні компоненти осідають першими(ядра, хлоропласти, мітохондрії)/. При підвищенні швидкості центрифуги осаджуються дрібніші органели (мікротоми, рибосоми). Швидкість седиментації кожного компоненту залежить від його розмірів і форми, і звичайно виражається коефіцієнтом седиментації – S на честь шведського фізика Теодора Сведберга, який розробив даний метод.
Для вивчення функціональної активності різних компонентів клітини іноді використовують методи мікрохірургії. Великі можливості у вивченні фізіології клітини відкривається з розвитком методів генної інженерії і біотехнології. Технології рекомбінантних ДНК у прикладних галузях дозволяють створювати транс генні організми з новими властивостями й ознаками, необхідними людині.
3. Еволюція клітини
За основу еволюційної теорії органічного світу приймається біохімічна гіпотеза російського біохіміка О.І.Опаріна (1922), відповідно до якої із простих неорганічних сполук – карбідів, окислів гірських порід, аморфного вуглецю і водню - могли створюватися первинні органічні речовини - можливо, вуглеводні. Завдяки радіації газовим розрядам, високій температурі і хімічним реакціям утворюються сполуки, подібні до ліпідів, амінокислот,нуклеїнових кислот. Сполуки ці накопичувалися як у земній атмосфері так і в океані. У воді вони створювали згустки в результаті об’єднання молекул і побудови інших комплексів. Такі згустки називають коацерватними краплями чи коацерватами. На границі між коацерватами і зовнішнім середовищем скупчувались молекули ліпідів, що привело до утворення примітивної клітинної мембрани. В міру росту коацервати могли розпадатись на дрібніші краплі, що було прообразом розмноження - більшість дрібних коацервантів також були здатні до вибіркової адсорбції.
Пізніше був розроблений лабораторний метод синтезу РНК у результаті проведення подібних експериментів. Це стало переломним моментом, тому, що полінуклеотиди здатні вже до якісно нової хімічної реакції – матричного синтезу, тобто до само подвоєння. Однак переконливо пояснити виникнення механізму, за допомогою РНК направляла б синтез білків, появу ДНК і здатність систем до самовідтворення, ця теорія не змогла.
В ході біохімічної еволюції утворилась величезна кількість складних органічних речовин. Так деякі клітини стали автотрофами. Поява автотрофів прискорила конкуренція гетеротрофів за їжу. Синтез складених органічних речовин різко зменшився, що зменшило кількість гетеротрофів, але одночасно підвищило стійкість процвітаючих форм життя - автотрофів. У таких умовах деякі анаероби вимирають, інші заповнюють екологічні ніші, практично позбавлені кисню. Треті вступають у симбіоз з аеробними клітинами і пізніше утворюють з ними міцну асоціацію.
Усі сучасні одноклітинні і багатоклітинні організми поділяються на дві групи – прокаріоти і еукаріоти. До прокаріот відносяться тільки бактерії і синьозелені водорості (ціанобактерії). До еукаріот – зелені рослини, гриби, слизовики і тварини.
4. Ультра структура (мікроскопічна будова)
Клітина – основна структурно - функціональна одиниця живої матерії, елементарна біологічна система всіх організмів, за винятком вірусів. У клітині здійснюються всі життєві процеси : живлення, генерація енергія виділення, новоутворення її елементів, розподіл,, реакція та подразнення і т.д. (рис.1.2).
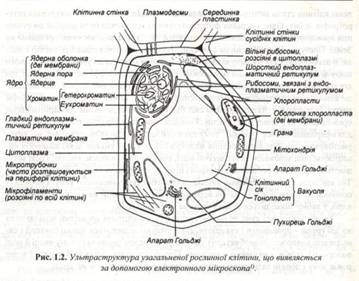
Поняття про клітину як складну відкриту біологічну систему постійно розвивається. Е.Ліберт (1976) вважає, що доцільно виділити три складові частини клітини : оболонку, протоплазму і вакуолю, а ряд структурних елементів протоплазми він створив ядро, мітохондрії і пластиди як носіїв генетичної інформації. Рибосоми, що містяться як у ядрі так і цитоплазмі. він виділяє окремо.
В.Г. Храновський (1982) пропонує поділяти клітину на протопласт і його продукти, а вакуолю розміщує між ними. Рибосоми знаходяться в комплексі цитоплазми.
С.І. Лебедєв (1988) розділяє клітину на дві основні частини – вміст (протопласт) і оболонку. Протопласт включає ядро і цитоплазму, у якій знаходяться органели. ВВ.Полєвой (1989), перелічивши складові частини клітини, не дотримується ніякої систематизації.
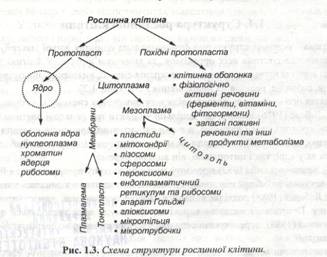
Узагальнюючи всі ці погляди, Є.М.Макрушина (1995) пропонує таку логічну схему структури рослинної клітини (рис.1.3).
5. Біологічні мембрани їх будови і функції
Структура і функції мембрани. Мембрани – пограничні цитоплазматичні структури, що забезпечують діалектичну єдність розмежування і зв’язку компонентів. До мембран відносяться ультротонкі структури, що оточують протопласт (пластична мембрана, плазма лема), вакуолі (тонопласт), ядро, пластиди, мітохондрії, апарат Гольджі, ендоплазматичний ретикулум, мікротоми й інші компоненти цитоплазми (рис. 1.4)..
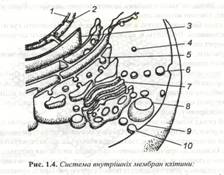
1- Ядерна мембрана, 2- ядерна пора, 3- ендоплазматична сітка, 4- мікротома, 5-перехідний міхурник, 6- везикула, 7- диктиосома, 8- тонопласт, 9- плазмолема, 10- екзоцитоз.
Існує самостійна наука – мемброналогія, що займається вивченням мембранних систем різного походження. Проблема мембран має велике значення не тільки для теорії, але й для прикладної біології.
Найважливішими функціями мембран є :
- забезпечення діалектної єдності розмежування і зв’язки клітинних компонентів.
- підтримка гомеостату – сталості складу середовища в кожному компртменті клітини.
- здійснення обміну речовин, енергії й інформації між клітиною і навколишнім середовищем.
- з мембранами зв’язана найважливіші біохімічні процеси клітини, оскільки в них локалізовані ферменти, що каталізують синтез, гідроліз,окислювання.
- на мембранах хлоропластів і мітохондрій здійснюються складні процеси біоенергетики, що забезпечують клітину енергією.
- мембрани виконують також рецепторно-регуляторну функцію : сприймаючі зовнішні і внутрішні подразнення і передаючи сигнали про них, забезпечують адаптивні відповіді клітин.
Основні компоненти мембран - ліпіди і білки. Мембрани мітохондрії відрізняються високим вмістом білків.
Мембрані білки поділяються на три групи :
1) Структурні білки, зв’язані з іншими компонентами мембран. Вони у звичайній мірі гідрофоюні, хімічно малоактивні.
2) Скорочувальні білки, що здатні в присутності АТФ змінювати густоту, в’язкість, світорозсіювання. Вважається, що вони здатні здійснювати активний транспорт речовин через мембрану.
3) Білки – ферменти, зв’язані з мембраною досить лабільно, що обумовлює зміну їх каталітичної активності в широкому діапазоні.
Хімічний склад мембран визначає їхні структурно-функціональні властивості. Особливе місце серед клітинних мембран займає плазматична мембрана. Ця периферична структура відмежує клітину від зовнішнього середовища і той же час забезпечує зв'язок з ним. Плазмалема є структурою, що здійснює вибіркове надходження речовин у клітину. а також визначає швидкість і спрямованість їх переміщення. Вона першою контактує з поживними й отруйними речовинами, фітогормонами і іншими сполуками, і, взаємодіючи з ними, чітка їх розпізнає.
В 1972 році С.Сінгер і Г.Ніколсон запропонували рідинно-мозаїчну модель структури біологічних мембран.(рис 1.5).
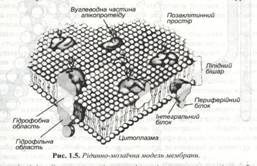
Відповідно до цієї моделі, основу мембран складає подвійний ліпідний шар, у якому присутні ліпіди трьох типів - фосфоліпіди, холестерол, і гліколіпіди. Усі вони відносяться до амфіпатичних молекул. Так, наприклад типова молекула фосфоліпіду має полярну головку (рис 1.6), основу якої складає гліцерил і два гідрофобних вуглеводневих хвости, представлені насиченими чи ненасиченими жирними кислотами.
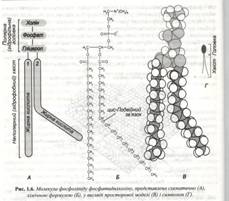
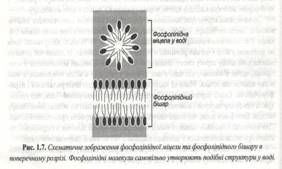
І одночасно з гліколіпідами у водному середовищі мимовільно утворюється бішар. При цьому гідрофобні хвости виявляються захованими й у контакті з водою залишаються тільки полярні головки. (рис. 1.7).
Схоже поводяться і молекули холестеролу, досить велику кількість, досить велику кількість якого містить саме плазматичний мембран. Взагалі, ліпідний склад внутрішнього і зовнішнього моно шарів мембрани різний. У плазматичній мембрані білки складають близько 50% маси. Як правило вони не розчинні у воді і погано розчинні в органічних розчинниках.
Білки часто називають відповідно інтегральними і периферичними. Серед перших широко представлені трансмембранні білки, що пронизують мембрану наскрізь і контактують з водним середовищем по обидва боки бішару ліпідів. Інші білки лише частково занурені в мембрану чи просто примикають до неї. У цілому білки утворюють на мембрані своєрідну мозаїку, іноді вільно переміщуючись по ліпідному бішару, а іноді й утримуючись на місці.
Транспорт речовин через мембран. Життєдіяльність організму, функціонування всіх його органів і систем можливі лише при постійному обміні речовин. Транспорт речовин повинен забезпечувати підтримку в клітині відповідного рівня рН і певної концентрації іонів, що сприяють ефективній роботі ферментів. У результаті транспорту всі тканини організму забезпечуються поживними речовинним,що служать джерелом енергії а також будівельним матеріалом для утворення клітинних компонентів.
Здатність клітин простояти змінам середовища і зберегти динамічну відносну складність складу називають гомеостазом. У рослині основну роль у підтримці гомеостазу грає транспорт речовин через плазма лему і тонопласт. Перша регулює приплив у клітині іонів і води з зовнішнього середовища і виділення баластових і надлишкових іонів, другий – надходження в протоплазму запасних субстратів із вакуоль при їх нестачі і видалення у вакуолю – при надлишку. Робота цих двох мембран також забезпечує стабілізацію осмотичного потенціалу клітини.
Відомі механізми пересування речовин через мембран поділяються на дві категорії – пасивний і активний транспорт. Пасивний транспорт – це рух речовин за законами дифузії й осмосу, який не вимагає витрат енергії. Дифузія – це переміщення молекул чи іонів за градієнтом концентрації: з області з вищою в область їз нижчою концентрацією. Осмос - це дифузія води через напівпроникну мембрану з області з низькою концентрацію розчинної речовини в область із високою.
Поряд з цими механізмами на характер дифузії впливає і загальний електричний градієнт мембрани. У принципі будь яка молекула шляхом дифузії може проникнути через ліпідний бішар (рис.1.8).
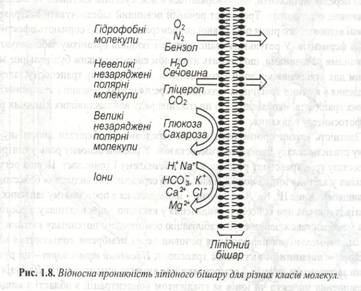
Набагато повільніше транспортуються великі полярні молекули (глюкоза,сахароза, амінокислоти). У цілому клітині мембрани проникні для різних полярних молекул. Відомо, що за їх перенос відповідальні специфічні білки. Такі транспортні білки присутні у всіх мембранах, але механізм їх роботи відрізняється певною специфічністю (рис. 1.9).
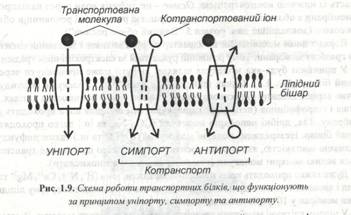
Одні з них просто переносять яку-небудь розчинену речовину з однієї сторони мембрани на іншу. Такий транспорт називають уніпортом. Інші функціонують як транспортні системи,у яких перенос однієї речовини залежить від переносу іншої речовини : або в тому ж напрямку (симпорт), або в протилежному (антипорт).
Деякі транспортні білки, що здійснюють пасивний транспорт, формують гідрофільні канали, які дозволяють окремим молекулам проходити бішар мембран за рахунок простої дифузії. Такі білки називаються каналоутворюючими (рис.1.10).
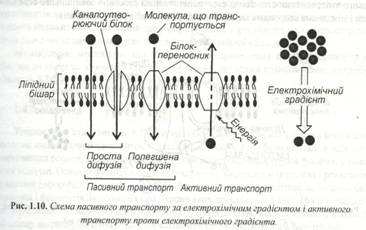
Інші білки здатні зв’язувати молекулу речовини, що транспортується, і переносити її через мембрану. Вона називається білками-переносниками, а весь процес – полегшеною дифузією.
Транспорт здійснюється проти електрохімічного градієнта, він сполучений зі споживанням енергії АТФ і називається активним. Механізми активного транспорту досить складні і до кінця не вивчені. У більшості клітин у плазматичній мембрані діє так званий калій-натрієвий насос. Активно діючий насос бере участь в осморегуляції клітин, а також шляхом сим порту чи анти порту разом з іонами викачує чи накачує цукри, амінокислоти і багато інших речовин. Внутрішній вміст клітини заряджений негативно стосовно зовнішнього середовища. Тому, відповідно до електричного градієнта, катіони прагнуть у клітину, а аніони відштовхуються. Концентрація іонів калію усередині клітини вища, ніж зовні. Для іонів натрію характерно зворотне співвідношення (рис.1.11).
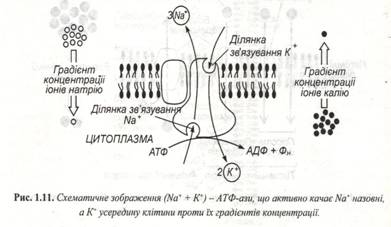
Висока концентрація калію необхідна кітлинам для білкового синтезу, гліколізу, фотосинтезу й інших життєво важливих процесів.
Калій-натрієвий насос характерний для клітин галофітів.У інших вищих рослин,грибів і водоростей у клітинах присутній протонний насос.У наслідок цього на мембрані генеруються електричний і концентратний потенціали. Перший забезпечує рух катіонів, а другий – дифузійний відтік протонів у клітину.
Транспортні білки опоседковують, проникнення через клітинні мембрани багатьох полярних молекул невеликого розміру, однак вони не здатні транспортувати макромолекули, наприклад полісахариди. При переносі макромолекул відбувається послідовне утворення і злиття оточених мембраною пухирців. Виділення із клітини зветься - екзоцитоз. Екзоцитоз характерний як для тваринних. так і для рослинних клітин. У клітині таким способом відбувається доставка вуглеводних компонентів до споруджуваної клітинної стінки. Процес, зворотній екзоцитозу, називають ендоцитозосм, але такий транспорт речовин у рослинних клітин спостерігається рідко через наявність у них твердої полісахаридної оболонки.
6.Цитоскелет, мікротрубочки і мікрофіломенти, будова і функції
Цитоплазма являє собою основну речовину клітини: цитоплазматичний матрикс (гіалоплазму) і мережу різних мембранних компонентів. Вона має вигляд своєрідних тривимірних решіток, побудованих з тонких білкових тяжів діаметром 3-6 нм, що заповнюють усю клітину. Усі компоненти цитоплазми виявляються немов підвішеними до цієї мікротрабекулярної решітки. Разом з мікротрубочками і мікрофіламентами вона складає так називний цитоскелет клітини. Мікротрубочки – це тонкі органели діаметром 24 нм, побудовані з білка тубуліну, що сягають у довжину кількох мікрометрів. При розподілі клітини вони складають основу веретена поділу і також беруть участь у внутрішньоклітинному транспорті. Мікрофіламенти – це nt; тонкы былковы нитки (дыаметром 507 нм), що складаються з актино- і міозиноподібних білків. Вони утворюють рухову систему клітини при рухах цитоплазми чи деяких структур всередині неї.
Цитоскелет поділяє клітку на дві фази : багату білком і багату водою, що заповнює простір між тяжами. Обидві фази мають консистенцію гелю чи золю (більш рідкий стан), що може регулюватися самою клітиною. При цьому цікаво відзначити, що присутність іонів двохвалентних металів підвищує активність деяких білків, які у свою чергу підсилюють полімеризацію білків цитоскелету. У тому числі формування поперечних зшивок між ними. Це виявляється за збільшенням в’язкості цитоплазми. Подібна структура гіалоплазми пояснює ще одну її важливу властивість - явище тиксотропії. Тобто гіалоплазма знаходиться на грані між розчинністю і нерозчинністю.
Гіалоплазма не тільки підтримує внутрішню структуру клітини. Тут же протікають тисячі біохімічних реакцій гліколізу, біосинтезу і розпаду цукрі, ліпідів, нуклеїнових кислот, білків, тощо.
7. Ядро, його будова і функції
Уперше ядро було описано і 1833 році Робертом Броуном, що знайшов його в цитоплазмі клітин тичинкових ниток традесканції. Згодом ядра були описані у всіх клітинах вищих рослин, за винятком зрілих ситоподібних трубок флоеми, у яких вони відсутні. Основні функції клітинного ядра – це збереження, передача і реалізація спадкової інформації, а також регуляція більшості процесів у клітині за допомогою гормонів і ферментів, інформація про будову яких знаходяться в ядрі. Носієм спадкової інформації, що передається в поколіннях, служить ДНК яка в основному входить до складу ядерної речовини будь – якої клітини, а також міститься й в цитоплазмі.
Клітини рослин бувають одноядерними, однак у деяких нижчих рослин можуть переважати двоядерні і багатоядерні клітини. Форми і розміри ядер коливаються не тільки у різних рослин, а також і в окремих тканинах однієї й тієї ж рослини. Зазвичай ядра мають кулясту чи яйцеподібну форму з діаметром від 10 до 30 мм. Клітинне ядро, що не поділяється, укладене в щільну ядерну оболонку, утворену двома мембранами, що розділені порожнинами шириною 20-40 нм. – перинуклеарним простором. (рис. 1.12).
Зовнішня ядерна мембрана складає єдине ціле з мембраною системою ендоплазматичної мережі і також як гранулярна ЕПС, вона із зовнішньої сторони часто буває засіяна функціонуючими рибосомами. В окремих ділянках ядерної оболонки, названих ядерними порами, зовнішня мембрана стуляється з внутрішньою ядерною мембраною. Пори особливо помітні на препаратах, отриманих методом заморожування – сколювання (рис. 1.13). Через них відбувається обмін різними речовинами між ядром і цитоплазмою.
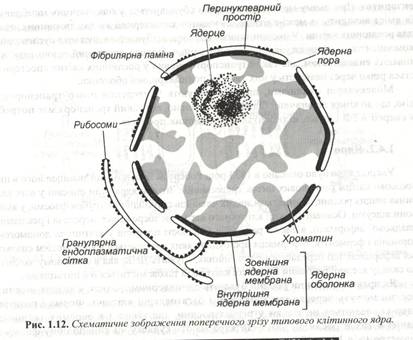
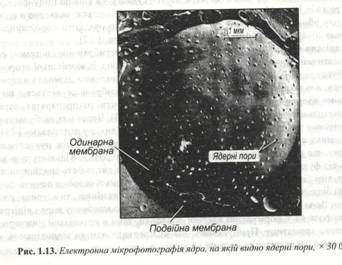
Кількість пор залежить від функціонального стану клітини – чим вища активність спадкоємного апарата, тим більше їх число. В області пор із внутрішньої сторони починається щільний білковий шар, що називається фібрилярною ламіною, чи ядерною пластиною. Вона на всьому протязі підсилає внутрішню мембрану ядра і відіграє ключову роль як у формуванні ядерної оболонки, так і організації нижче розташованого хроматину. Припускають, що поліпептиди ламіни відповідають за руйнування і реорганізацію ядерної оболонки під час мітозу.
Вміст ядра являє собою гелеподібний матрикс, називаний ядерним соком чи нуклеолплазмою. Вона містить різні хімічні речовини - іони, білки, ферменти, і нуклеотиди у вигляді колоїдного чи справжнього розчину. У неклеоплазмі розташовується хроматин, що складається з комплексу ДНК. РНК і білків гістонів. Гістони об’єднані з ДНК у структури, що нагадують намистини (нуклеосоми) і виконують структуру і регуляторну роль. Під час поділу клітини хроматин піддається конденсації з утворенням туго скручених, спаралізованих ниток – хромосом. Сукупність хромосом у клітині називають хромосомним набором. Розділяють два типи наборів – гаплоїдний і диплоїдний. Гаплоїдний набір за числом хромосом удвічі менший диплоїдного і типовий для генеративних клітин і гаметофіту. Диплоїдний набір складається з двох гаплоїдних наборів материнського і батьківського організмів і міститься у всіх соматичних клітинах рослин і тварин.
У неоплазмі ядер різних клітин може знаходитися одне чи кілька ядерець. На відміну від цитоплазматичних органел, ядерце не має мембрани й утворюється на певних ділянках молекул ДНК – ядерцевих організаторах, на яких знаходиться велике число генів, що кодують рибосомальну РНК. Так же відбувається її синтез і з’єднання з білками при формуванні великих і малих субодиниць рибосом. Надалі субодиниці через ядерні пори залишають ядро і виходять у цитоплазму, де відбувається їх остаточна збірка у функціонально зрілі рибосоми.
8. Ендоплазматична сітка, будова і функції
Одним з важливих відкриттів, зроблених за допомогою електронного мікроскопа, було виявлення в 1945 р. К.Портером складної системи мембран, що пронизує цитоплазму всіх еукаріотичних клітин. Ця мережа мембран, що одержала назву ендоплазматична сітка чи ендоплазматичний ретикулум. Являє собою систему субмікроскопічних канальців, трубочок, округлих пухирців і плоских мішечків, обмежених одинарною мембраною. Вважають, що хоча мембрана ЕПС має числені складки і вигини, вона утворює одну, практично безперервну поверхню, що обмежує єдиний замкнутий мішок. (рис. 1.14).
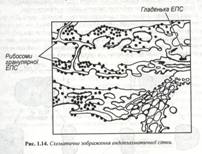
Цей внутрішній простір, який називають порожниною ЕПС, часто займає близько 10-16 % загального обсягу клітини. Мембрани ЕПС складають єдине ціле з зовнішньою ядерною мембраною, але не зв’язані з плазмалемою (рис. 1.15). У клітинах можна виділити дві функціонально різні області ЕПС : шорсткувата (гранулярна) ЕПС, мембрана якої, так само як і зовнішня мембрана ядра, усіяна рибосомами, розташованими на зверненій до цитоплазми стороні мембрани, і гладенька (гранулярна) ЕПС, позбавлена рибосом. Ці дві області значно розрізняються і за формою: шорсткувата ЕПС являє собою стінки сплющених мішечків, які називають цистернами, а гладеньку НПС складається з мережі тонких трубочок. У гранулярній мережі синтезуються різні білки. Серед них – мембранні білки, у тому числі й глюкопротеїди, ферменти, необхідні для синтезу полісахаридних клітинах стінок та інші. Транспортуючись по цистернах, деякі з цих сполук доставляються в апарат Гольджі, відтіля вони виходять назовні або надходять в інші органели чи відкладаються у вигляді запасних гранул. Інші білкові речовини після синтезу виділяються в цитозоль для використання в цій же клітині.
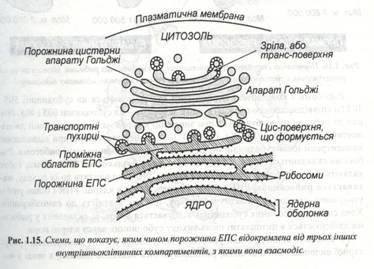
По канальцях і трубочках ЕПС здійснюється не тільки транспорт речовин усередині клітини. Вона бере участь також у міжклітинних взаємодіях через плазмодесми, зв’язуючи цитоплазми клітин у єдину систему - симпласт.
9. Рибосоми, будова і функції
Рибосомами називають сферичні чи грибоподібні, дуже дрібні органели діаметром близько 22 нм. Вони були вперше описані і 1953 році американським біологом Д.Е.Паладе, який довів, що рибосоми являють собою нуклеопротеїди.
Кожна рибосома складається з двох субодиниць – великої і малої. Для характеристики рибосом використовують їхню властивість, як і всяких часток і молекул, осаджуватися з постійною швидкістю під дією відцентрової сили при диференційованому центрифугуванні. Досліди по седиментації виявили існування двох головних типів рибосом, що були названі 70S і 80S – рибосомами. Перший тип являється у прокаріот, а більші 80 S –рибосомами – в еукаріотичних клітинах (ри.с.1.16).
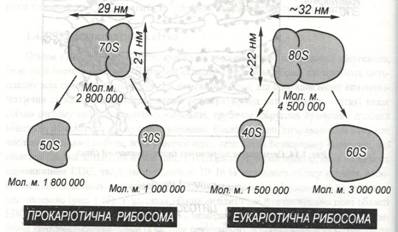
Рис. 1.16. Порівняння структур прокаріотичних і еукаріотичних рибосом. Незважаючи на розходження в структурі, ці рибосоми дуже подібні у функціональному відношенні.
Рибосоми з константою седиментації 70 S поділяють на субодиниці 50S і 30S, а 80S – рибосоми - на субодиниці 60S і 40S. Розпад на субодиниці відбувається при зниженні в середовищі концентрації двовалентних іонів металів (Mg, Ca, Co, Mn). Цей процес зворотний : при відновленні концентрації іонів субодиниці об’єднуються, утворюючи нативні рибосоми. Рибосоми складаються з приблизно рівних за масою кількістю РНК і білка, тобто являють собою рибонуклеопротеїнові частки. РНК, що входить до їх складу, називається рибосомною РНК (рРНК) і синтезується в ядерці.
У клітинах вищих рослин рибосоми звичайно зв’язані з мембранами гранулярної ендоплазматичної сутки, розташовуючись безладно лінійно, а іноді у вигляді розеток чи спіралей на їх зовнішній поверхні. У різних рослинних клітинах число рибосом коливається від декількох тисяч до декількох десятків тисяч одиниць. Їх концентрація у клітинах і тканинах може змінюватися у процесі онтогенезу і залежить від живлення, водного режиму, температури й інших умов, а також функціональної активності клітини.
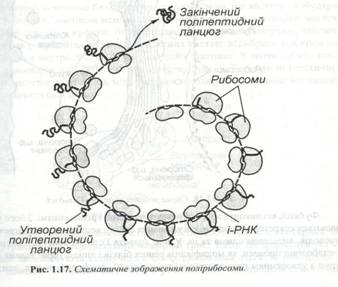
Установлено, що на рибосомах відбувається біосинтез білка, тобто зв’язування активованих амінокислот і укладення їх у поліпептидний ланцюг відповідно до генетичної інформації, отриманої з ядра через інформаційну РНК. На поверхні іРНК відбувається взаємодія між комплексами « амінокислота – транспортна РНК» і самою рибосомою. З рибосомами одночасно можуть з’єднуватися тільки два комплекси і замикається тільки один пектиновий зв'язок. Потім рибосома переміщується по іРНК, до неї підходить наступний комплекс «амінокислота – тРНК» і замикається новий пептидний зв'язок. Так продовжується до закінчення синтезу поліпептидного ланцюга. Цей процес називається трансляцією. Рибосоми діють в основному не ізольовано, а спільно й упорядковано. Угрупування рибосом, що транслює іРНК і синтезує той самий білок, одержало назву полісоми. В одній полісомі може знаходитися від 5 до 70 рибосом. На ЕПС полісоми виявляються у вигляді характерних завитків (рис.1.17).
10. Мітохондрії, будова і функції
У рослин мітохондрії були вперше виявлені в 1904 р. Ф.Мевесом в клітинах пиляків латаття. Вони являють собою округлі чи гантелеподібні тіла, розміри яких надзвичайно мінливі і значною мірою залежать від функціонального стану у клітин, осмотичного тиску і рН середовища. Мітохондрії збільшуються в гіпотонічних речовинах і зменшуються в гіпертонічних, а в кислому середовищі набувають пухирчастої форми. Товщина мітохондрії постійна (близько 1.5 мкм), у той час як довжина помітно коливається, досягаючи 7-10 мкм і більше.
Мітохондрії обмежені двома мембранами (рис. 1.19).
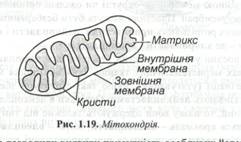
Міжмембранний ростір, чи перимітохондріальний простір, заповнений основною безструктурною речовиною, що містить глобулярні білки і деякі ферменти. Зовнішня мембрана гладенька, а внутрішня утворює численні гребнеподібні складки - кристи. Вони істотно збільшують її поверхню, забезпечуючи площу для розміщення мультиферментиних систем. Мембрани відокремлюють від цитоплазми внутрішній вміст мітохондрій - матрикс. У матриксі містяться рибосоми і мітохондріальна ДНК, що має кільцеву будову.
Сучасні методи дозволили виявляти присутність особливих «елементарних часток» на внутрішній мітохондріальній мембрані (рис.1.20). Це ферменти АТФ - синтетази, що забезпечують сполучення фосфорилювання АДФ із реакціями в дихальному ланцюзі.
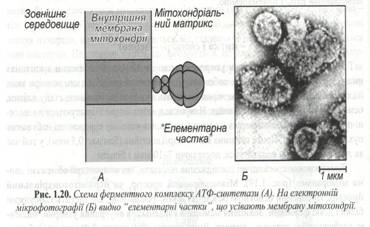
В основі цих часток розташовані компоненти самого дихального ланцюга. Таким чином, основна задача мітохондрій – це синтез АТФ у результаті циклічного окислювання ди- і три карбонових кислот і аеробних реакцій електронотранспортного ланцюга дихання.
Кількість мітохондрій варіює від десятків до десятків тисяч на клітину, змінюючись в онтогенезі, отже вона визначається рівнем метаболізму. У залежності від ділянки клітини, де необхідні електричні витрати, мітохондрії з течією цитоплазми переміщуються в ту чи іншу ділянку.
11. Пластиди, будова і функції
Перші описи пластидів були зроблені в 1676 р. Антоні ван Левенгуком, але детальні дослідження цих структур провів Шимпер в 1882 р.
Система органоїдів, названих пластидами, типова для клітин рослин. Це обмежені подвійною мембраною округлі чи овальні органоїди, що містять внутрішню систему мембран. Пластиди можуть бути безбарвними чи забарвленими. Серед безбарвних розрізняють про пластиди, лейкопласти і етіопласти. Забарвлені пластиди – це хлоропласти і хромопласти.
У вищих рослин усі типи пластид утворюються з про пластид, що є присутніми у клітинах меристематичних тканин. Їхня внутрішня мембрана має лише невеликі інвагінації (утягування). Якщо такі структури зберігаються в зрілих клітинах, їх називають лейкопластами. Оскільки в лейкопластах відкладаються запасні речовини, то назву вони одержують залежно від типу запасної сполуки : якщо запасається крохмаль – амінопласти, жири – олеопласти, білки - протеїнопласти. Етіопласти утворюються при вирощуванні рослин у темряві. При освітлені вони перетворюються в хлоропласти.
Хлоропласти - зелені пластиди рослин. У них здійснюється – фотосинтез. Це овальні тіла довжиною 5-10 мкм і 2-3 мкм у діаметрі (рис.1.21).
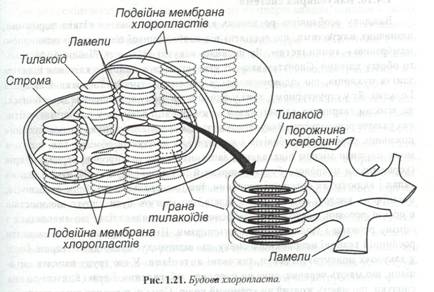
Внутрішня мембрана утворює безліч плоских, заповнених рідиною мішків, так званих тилакоїдів. Декілька таких телакоїдів формують купку - грану. Окремі грани хєднані довгими поодинокими тилакоїдами – ламелами. На мембранаї тилакоїдів локалізовані зелені пігменти – хлорофіли, а також жовті і червоні – каротиноїди. На мембранах тилакоїдів відбуваються світові реакції фотосинтезу. Внутрішнє, гомогенне середовище хлоропластів називається стромою. У ній знаходяться розчинні ферменти, що здійснюють темнові реакції фотосинтезу. Надлишок вуглеводів, що утворюється в процесі цих реакцій, може запасатися у вигляді зерен крохмалю. З мембран часто бувають зв’язані кулясті краплі ліпідів, у яких акумулюються ліпідні компоненти мембран. Кільцева ДНК і рибосоми дозволяють пластидам, так само як і мітохондріям, ділитися незалежно від поділів самої клітини. Іноді кількість хлоропластів може збільшуватися не шляхом поділу, а брунькуванням.
Хромосоми містять у стромі велику кількість каротиноїдів. Пігменти, що розташовані не на мембранах і знаходяться в атмосферному стані, часто представлені у вигляді кристалів. Присутністю каротиноїдів пояснюється забарвлення плодів томата, горобини, осіннього листя, коли хлорофіл у них руйнується і виявляється колір інших пігментів.
12. Мікросоми
Уперше були виявлені в 1880 р. Ганштейном. Мікросоми являють собою кулясті тіла діаметром від 0.2 до 1.5 мкм, обмежені елементарною мембраною. Первинні мікросоми утворюються за рахунок діяльності ендоплазматичної сітки. У клітинах рослин виявлені два типи мікросом, що виконують різні функції - пероксисоми і гліоксисоми.
Пероксисоми численні в клітинах листків, де вони тісно зв’язані з хлоропластами. У них відбувається окислювання синтезованої в процесі фотосинтезу гліколевої кислоти з утворенням амінокислоти гліцину, яка вже в мітохондріях перетворюється в серин. При роботі ферментних систем пероксисом утворюється пероксид водню, який руйнується в цих органоїдах ферментом каталазою, що міститься в органоїдах.
Гліоксисоми з’являються при проростанні насіння, у якому запас поживаних речовин представлений здебільшого жирами. Вони містять ряд ферментів, необхідних для перетворення жирів у цукри в процесі окислювання жирних кислот і реакцій гліоксилатного циклу.
13. Вакуолярна система
Важливу особливість рослинних клітин мають одна чи кілька порожнин, називають вакуолями, що відділені від навколишньої цитоплазми одинарною мембраною – тонопластом. Як правило, вакуолі займає більше 50% всього обсягу клітини. Спочатку вакуолі утворюються в молодих клітинах шляхом злиття пухирців, що відокремлюються від ендоплазматичної сітки й апарата Гольджі. Як у структурному, так і у функціональному відношеннях вони близькі до лізосом тваринних клітин. Разом з тим функції вакуоль у рослинних клітинах разюче різноманітні. Вакуолі служать для транспортування і нагромадженні поживних речовин, метаболітів і непотрібних продуктів обміну. Вакуоле може служити місцем відкладання запасних білків, цукрі. Вакуолярний сік має складний склад і включає як органічні речовини, так і мінеральні солі. Крім вуглеводів, органічних кислот, амінокислот і білків, що можуть бути удруге використані в обміні речовин, вакуолі містять феноли, таніни, алколодїи, що виводяться з обміну речовин і ізолюються від цитоплазми. Ці сполуки можуть надавати рослинній тканині неприємного смаку, що відштовхує травоїдних тварин. Іноді у вакуолях присутні пігменти, так звані антоціани. У цю групу входять антоціани, що мають червоне, синє чи пурпурне забарвлення, і деякі близькі до них сполуки, що мають жовтий чи кремовий колір. Саме ці пігменти визначають забарвлення квітів у рожевих фіалок, жоржин та ін..
Вакуолярний сік може містити ряд гідролітичних ферментів, і тоді при житті клітини вакуолі діють як лізосоми,здійснюючи автофагію – знищення непотрібних клітині структур з утворенням окремих автофагічних вакуоль.. Після загибелі клітин тонопласт як і всі мембрани утрачає свою вибірну проникність і вже не здатний виконувати бар’єрну роль.
Одна з головних функцій вакуолі – це участь в осмотичних явищах. Вода звичайно надходить у концентрований клітинний сік шляхом осмосу через вибірково проникний тонопласт. У результаті в клітині розвивається тургорний тиск і цитоплазма притискається до клітинної стінки. Такий тиск – необхідна умова росту клітин розтяганням, причому обсяг клітин збільшується без збільшення обсягу самої цитоплазми.
14. Апарат Гольджі, будова і функції
Структуру, відому тепер як апарат Гольджі уперше знайшов у тваринних клітинах у 1898 р. італійський гістолог К. Гольджі. Однак у рослинних клітках існування подібних структур було достовірно встановлене в результаті робіт із застосуванням електронного мікроскопа Бюва і Портером лише в 1957 році. Апарат Гольджі. Являє собою купку сплощених мембранних мішечків, так званих цистерн, і зв’язану з ними систему пухирців, які називають пухирцями Гольджі, чи везикулами. Ряд окремих купок називають диктіосомами. (рис. 1.18).
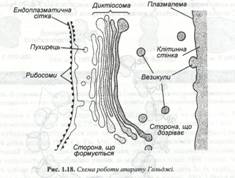
У зоні диктіосоми розрізняють дві ділянки. З однієї сторони шляхом злиття пухирців, що відбруньковуються від гладенької ЕПС, постійно утворюються нові цистерни. Ця сторона купки, що формується, опукла, тоді як інша, де завершується дозрівання і цистерна знову розпадаються на пухирці, має увігнуту форму. Локалізація, величина і розвиток елементів змінюється залежно від типу клітини і її фізіологічного стану. Так, АГ найбільш розвинутий в активно функціонуючих клітинах у процесі старіння він поступово атрофується, а потім зникає.
Функції, які виконує апарат Гольджі, чисельні та різноманітні. З його участю здійснюються секторні процеси, регулювання вмісту води в клітині, нагромадження вуглеводів, виділення слизів та ін.. У зоні апарата Гольджі можуть здійснюватися такі метаболічні процеси, як модифікація різних білків і ліпідів, фосфорилювання, ацилування амінокислот тощо. У рослинних клітинах вдалося простежити участь АГ в утворенні серединної пластинки і росту клітинних стінок. Установлено, що за допомогою везикул вуглеводні компоненти постачаються до плазми, при цьому транспорт у певному напрямку здійснюють мікротрубочки ;мембрана пухирців відбувається у плазмалему, сприяючи її росту і відновленню, а секретизовані рослини виявляються в клітинній стінці, де за допомогою ферментів синтезуються целюлозні мікрофібрини.
Таким чином, мембрання елементи апарата Гольджі є сполученою ланкою між мембранами ЕПС і плазмалемою.
15. Будова і функції клітинних стінок
Оболонку клітин, що поділяють і ростуть, називають первинною стінкою. Вона складається з целюлозних мікрофібрил, занурених у матрикс (рис. 1.22).

Мікрофібрили являють собою пучки довгих тонких молекул целюлози. До складу матрикса входять полісахариди і білки, багато з яких є ферментами, що каналізують синтез речовин, які входять до складу оболонки. Полісахариди матриксу представлені пектиновими сполуками і геміцелюлозами - змішаною групою полімерів занози, ксилоли й інших моносахаридів. Стінки сусідніх клітин з’єднує серединна пластинка, що складається з клейких желеподібних пектитів кальцію і магнію (рис. 1.23).
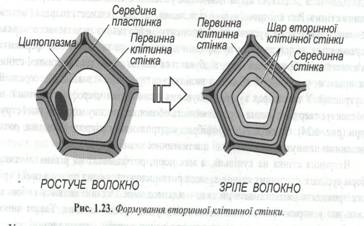
Клітинні стінки гідратовані – 60-70% їхньої маси звичайно складає вода. Разом з міжклітинниками вони складають так званий вільний простір - апопласт. По апопласту відбувається переміщення води і розчинених у ній речовин. Частка апопласту адсорбує різні іони, полегшує їхній подальший рух у клітину. У деяких клітин, наприклад, у клітин мезофілу листка, протягом усього життя зберігається тільки первинна стінка. У більшості клітин після закінчення росту на внутрішню поверхню первинної клітинної стінки відкладаються додаткові шари целюлози, унаслідок чого виникає вторинна клітинна стінка. Ріст клітинної стінки можливий завдяки апарата Гольджі. На вторинній оболонці виділяється три шари – зовнішній, середній і внутрішній. У цих шарах кут розташування целюлозних мікрофібрил різний, що забезпечує твердість, незначну розтяжність оболонки і високу міцність усієї структури. (рис. 1.24.) Орієнтація мікрофібрил контролюється мікротрубочками, розташованими на внутрішній поверхні плазматичної мембрани.

Вторинна стінка не суцільна, а має пори, розташовані на різних ділянках. Пори сусідніх клітинних стінок завжди розташовуються одна проти одної і крізь них проходять цитоплазматичні тяжі – плазмодесми. Останні являють собою канали, що утворюються з плазма леми і з’єднують сусідні клітини. Таким чином, протопласти окремих клітин з’єднуються в єдину систему - симпласт.
Вторинна стінка, що виникає в клітинах диференційованих тканин, поділяється значним структурним і хімічним змінам. Трахеальні елементи ксилеми і клітинами склеренхіми піддаються інтенсивній лігніфікації. Це відбувається при просочуванні шарів целюлози лігніном – складною полімерною речовиною, що відноситься до полі фенолів. Лігнін скріплює целюлозні волокна й утримує їх у єдиній структурі. Характер і ступінь одеревіння можуть бути різними. Лігніфікація характерна для механічних і провідних тканин.
У вторинних покривних тканинах відбувається опробування клітинних стінок. При цьому у вторинній клітинній стінці відкладається суберин – жироподібна речовина, до складу якої входять жирні кислоти і їх ефіри з гліцерином та іншими спиртами. Опробковані клітини широко відмирають і перетворюються у захисний шар. Суберин надає їм непроникності щодо води, газів, грибної та мікробної інфекції.
Зовнішні клітинні стінки епідермальних тканин деяких видів рослин можуть бути покриті кутикулою - тонкою плівкою, що складається з кутину (суміш високомолекулярних жирних кислот і їх ефірів). Часто кутикула покривається шаром воску, товщина якого у деяких рослин досягає 5 мм. Епідермальні тканини можуть піддаватися також ослизненню, тобто покриваються сумішшю високомолекулярних вуглеводів, білків, ферментів, і органічних кислот. Кутинізація й ослизнення захищають рослин від перегріву і надмірної втрати води.
Література
1. Грин Н., Стаут У.,Тейлор Д., Биология, Мир 1990.
2. Лебедев С.И. Физиология растений, Колос 1988.
3. Мусієнко М.М. Фізіологія рослин. Київ. Фітосоціоцентр, 2001.
4. Макрушин М.М., Макрушина Є.М., Петерсон Н.В., Мельников М.М. Фізіологія рослин. / За редакцією професора М.М.Макрушина Віниця: Нова Книга, 2006.
5. Полевой В.В. Физиология растений. Выссшая школа, 1989.
Перепечатка материалов без ссылки на наш сайт запрещена