Реферат: Зрительные пигменты
Реферат: Зрительные пигменты
Зрительные пигменты
Содержание
Зрительные пигменты
Поглощение света зрительными пигментами
Строение родопсина
Колбочки и цветовое зрение
Цветовая слепота
Свойства каналов фоторецептора
Молекулярная структура цГМФ-управляемых каналов
Передача сигнала в фоторецепторах
Метаболический каскад циклического ГМФ
Рецепторы позвоночных, деполяризующиеся при действии света
Усиление сигнала в каскаде цГМФ
Сигналы в ответ на одиночные кванты света
Литература
Зрительные пигменты
Зрительные пигменты сконцентрированы в мембранах наружных сегментов. Каждая палочка содержит около 108 молекул пигмента. Они организованы в несколько сотен дискретных дисков (около 750 в палочке обезьян), которые не связаны с наружной мембраной. В колбочках пигмент расположен в особых пигментных складках, которые являются продолжением наружной клеточной мембраны фоторецептора. Молекулы пигмента составляют около 80% всех белков диска. Зрительные пигменты настолько плотно упакованы в мембранах наружного сегмента, что расстояние между двумя молекулами зрительного пигмента в палочке не превышает 10 нм. Такая плотная упаковка повышает вероятность того, что фотон света при прохождении слоя фоторецепторных клеток будет уловлен. Возникает следующий вопрос: каким образом возникают сигналы при поглощении света зрительными пигментами?
Поглощение света зрительными пигментами
События, происходящие при поглощении света пигментом палочек - родопсином, изучались при помощи психофизиологических, биохимических и молекулярных методик. Молекула зрительного пигмента состоит из двух компонентов: белковой, называемой опсином, и хромофора, 11-цис-витамин А-альдегида, называемого ретиналем (рис.1). Следует уточнить, что хромофор содержит химическую группу, придающую цвет соединению. Количественные характеристики поглощающей способности пигментов были изучены при помощи спектрофотометрии. При освещении родопсина - зрительного пигмента палочек - светом разной длины волны, сине-зеленый свет с длиной волны около 500 нм поглощался лучше всего. Подобный результат был получен и при освещении отдельной палочки под микроскопом пучками света с разной длиной волн. Была выявлена интересная зависимость между спектром поглощения родопсина и нашим восприятием сумеречного света. Количественные психофизические исследования, выполненные на человеке, показали, что голубовато-зеленый дневной свет с длиной волны около 500 нм оптимален для восприятия сумеречного света в темноте. Днем, когда палочки неактивны и используются только колбочки, мы наиболее чувствительны к красному цвету, соответствующему спектру поглощения колбочек (об этом мы поговорим далее).
При поглощении родопсином одного фотона ретиналь претерпевает фотоизомеризацию и переходит из 11-цис в транс-конфигурацию. Этот переход происходит очень быстро: примерно за 10--12 секунд. После этого белковая часть пигмента также претерпевает серию трансформационных изменений, с образованием ряда промежуточных продуктов. Одна из конформаций протеиновой части - метародопсин II - наиболее важна для передачи сигнала (мы обсудим это далее в этой главе). На рис.2 показана последовательность событий при обесцвечивании и регенерации активного родопсина. Метародопсин II образуется уже через 1 мс. Регенерация пигмента после его распада происходит медленно, в течение нескольких минут; для этого необходимо транспортирование ретиналя из фоторецепторов в пигментный эпителий.
Строение родопсина
На молекулярном уровне белок опсин состоит из 348 аминокислотных остатков, образующий 7 гидрофобных зон, каждая из которых состоит из 20-25 аминокислот, составляя 7 трансмембранных спиралей. Ν-конец молекулы расположен во внеклеточном пространстве (т.е. внутри диска палочки), а С-конец находится в цитоплазме.

Рис.1. Структура родопсина позвоночных, встроенного в мембрану фоторецептора. Спираль несколько развернута, чтобы показать расположение ретиналя (указано черным). С - С-конец, N - N-конец.
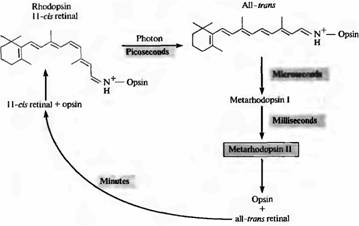
Рис.2. Выцветание родопсина на свету. В темноте 11-цис-ретиналь прочно связан с белком опсином. Захват фотона приводит к изомеризации all цис ре тиналя в троне ретиналь. При этом комплекс опсин all-тронс-ретиналь быстро превращается в метародо псин II, который диссоциирует на опсин и all троне ретиналь. Регенерация родопсина зависит от взаимодействия фоторецепторов и клеток пигментного зпителия. Метародопсин II включает и поддерживает в активном состоянии систему вторичных посредников.
Ретиналь соединен с опсином через остаток лизина, расположенный в седьмом трансмембранном сегменте. Опсин принадлежит к семейству белков, имеющих 7 трансмембранных доменов, в которое входят и метаботропные рецепторы медиаторов, такие как адренергические и мускариновые рецепторы. Как и родопсин, эти рецепторы передают сигнал ко вторичным посредникам посредством активации G-белка. Родопсин удивительно стоек в темноте. Байор подсчитал, что для спонтанной тепловой изомеризации молекулы родопсина необходимо около 3000 лет, или в 1023 больше, чем для фотоизомеризации.
Колбочки и цветовое зрение
Удивительные исследования и эксперименты, выполненные Янгом и Гельмгольцем в XIX веке, привлекли внимание к очень важному вопросу о цветном зрении, и сами же ученые дали четкое и точное объяснение этому феномену. Их вывод о существовании трех различных типов цветовых фоторецепторов выдержал испытание временем и был в последующем подтвержден на молекулярном уровне. Вновь можно процитировать Гельмгольца, который сравнил восприятие света и звука, цвета и звукового тона. Можно позавидовать ясности, силе и красоте его мысли, особенно в сравнении со сбивающими с толку виталистистическими концепциями, широко распространенными в XIX веке:
Все различия е цветовых тонах зависят от комбинации е различных пропорциях трех основных цветов... красного, зеленого и фиолетового... Подобно тому, как восприятие нани солнечного света и его теплоты зависит... от того, попадают ли лучи солнца на нервы, идущие от рецепторов зрения или от рецепторов тепловой чувствительности. Как предположил Янг в своей гипотезе, различие в восприятии различных цветов зависит просто от того, какой из 3 типов фоторецепторов больше активируется этим светом. Когда все три типа в равной степени возбуждены, получается белый цвет...
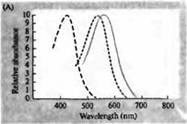
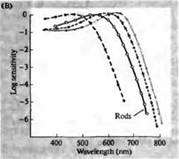
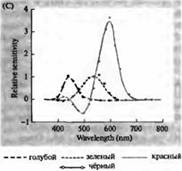
Рис. 3. Спектры чувствительности фоторецепторов человека и различных зрительных пигментов. (А) Кривые спектров чувствительности трех цветовых зрительных пигментов, показывающие пики поглощения на длинах волн, соответствующих голубому, зеленому и красному. (В) Спектры чувствительности колбочек к голубому, зеленому и красному цветам, и палочек (показано черным) у макак. Ответы регистрировались при помощи всасывающих электродов, усреднялись и нормализовывались. Кривые спектра палочки были получены при исследовании зрительных пигментов на людях. (С) Сравнение спектров колбочек обезьян и человека при помощи теста чувствительности к цвету. Непрерывная кривая показывает эксперимент по определению чувствительности к цвету у человека, при предъявлении ему света разной длины волны. Пунктиром показаны результаты, предсказанные на основе регистрации токов в отдельных колбочках, после коррекции поглощения света в хрусталике и пигментами на пути к наружному сегменту. Совпадение между результатами обоих опытов удивительно высокое.
Если мы проецируем на белый экран два пучка света различных цветов одновременно... мы видим только один цвет, более или менее отличный от обоих этих цветов. Мы сможем лучше понять выдающийся факт того, что мы способны воспринимать все оттенки в составе внешнего света путем смеси трех основных цветов, если мы сравним глаз сухом... В случае звука... мы слышим более длинные волны как низкие тона, а короткие волны - как высокие и пронзительные, кроме этого ухо способно улавливать одновременно много звуковых волн, т.е. много нот. Однако они β данном случае не сливаются в один сложный аккорд, подобно тому, как различные цвета... сливаются в один сложный цвет. Глаз не может показать разницу, если мы заменяем оранжевый цвет на красный или желтый; но если мы слышим ноты до и ми, звучащие одновременно, нам подобное звучание не кажется нотой ре. Если бы ухо воспринимало музыкальные тона подобно тому, как глаз воспринимает цвета, каждый аккорд мог бы быть представлен комбинацией трех постоянных нот, одной очень низкой, одной очень высокой и одной промежуточной, вызывая все возможные музыкальные эффекты только путем изменений относительной громкости этих трех нот... Однако мы способны видеть плавный переход цветов одного в другой через бесконечное множество оттенков и градаций... То, каким образом мы воспринимаем каждый из цветов... зависит е основном от строения нашей нервной системы. Надо признаться, в настоящее время ни у человека, ни у четвероногих не описана анатомическая база для подтверждения теории восприятия цвета.
Эти точные и дальновидные предсказания были подтверждены серией различных наблюдений. При помощи спектрофотометрии Вальд, Браук, МакНикол и Дартнэл с коллегами показали наличие в сетчатке человека трех типов колбочек с различными пигментами. Также Бейлор с коллегами сумели отвести токи от колбочек обезьян и человека. Было обнаружено, что три популяции колбочек имеют различные, но перекрывающиеся диапазоны чувствительности к голубой, зеленой и красной части спектра. Оптимальные длины волн для возбуждения электрических сигналов в точности совпали с пиками поглощения света зрительными пигментами, установленными при помощи спектрофотометрии и при психофизических экспериментах по измерению чувствительности глаза к цветовому спектру. В конечном итоге Натаисом были клонированы и секвенированы гены, кодирующие пигмент опсин в трех типах колбочек, чувствительных к красному, зеленому и голубому спектру.
Каким же образом молекулы различных зрительных пигментов способны предпочтительно улавливать свет определенной длины волны? Оказывается, родопсин - зрительный пигмент палочек и все три зрительных пигмента колбочек содержат в своем составе один и тот же хромофор, 11-цис-ретиналь. Однако аминокислотные последовательности белковой части пигмента отличаются друг от друга. Различиями всего в нескольких аминокислотах и объясняется различная их чувствительность к спектру.
Цветовая слепота
Хотя один тип фоторецептора не способен сам по себе воспринимать цвет, три типа колбочек, как показано на рис. 4, уже способны.
В принципе, двух типов колбочек с различными пигментами было бы достаточно для распознавания цвета, однако в этом случае ряд комбинаций длин волн воспринимался бы одинаково. Подобная ситуация возникает в случае, когда человек страдает цветовой слепотой. У таких людей, как было показано Натансом, имеются генетические дефекты, приводящие к отсутствию одного из пигментов. С высоты нынешнего состояния науки мы не можем не поразиться тому, как красиво молекулярные механизмы подтверждают блистательные и на удивление точные размышления Янга и Гельмгольца.
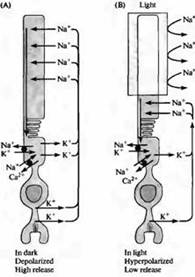
Рис. 4. "Темновой" ток в палочке. (А) В темноте ионы натрия проходят через катионные каналы в наружном сегменте палочек, вызывая деполяризацию; ионы кальция также способны проходить через зти катионные каналы. Петля тока проходит через перешеек палочки за счет того, что калиевый ток течет в направлении наружу во внутреннем сегменте мембраны. (В) Когда происходит освещение наружного сегмента, каналы закрываются из-за уменьшения внутриклеточной концентрации цГМФ, и палочка гиперполяризуется. Гиперполяризация проводит к уменьшению высвобождения медиатора. Концентрации натрия, калия и кальция поддерживаются внутри палочки специальными насосами и ионообменниками, расположенными в области внутреннего сегмента (черные кружки). Транпортеры кальция также находятся и в наружном сегменте.
Их идея о том, что основные атрибуты цветного зрения и цветовой слепоты должны быть обнаружены в самих фоторецепторах, были подтверждены при помощи прямых физиологических измерений, а также при изучении различий в строение пигментов на генетическом и белковом уровнях ный потенциал составляет порядка - 40 мВ, что далеко от равновесного калиевого потенциала ЕК, составляющего - 80 мВ.
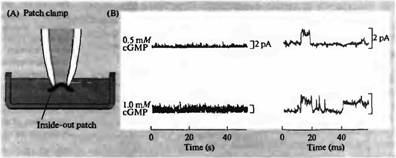
Рис. 5. Роль цГМФ в открытии натриевых каналов наружного сегмента палочек. Проводилась регистрация активности одиночных каналов при помощи inside-out patch clamp, находящихся в растворе с разной концентрацией цГМФ. Открытие канала приводит к отклонению тока вверх. Частота открытия каналов была очень низка в контроле, добавление цГМФ приводило к увеличению частоты открытия одиночных каналов, которая прямо зависела от концентрации цГМФ
Входящий ток в темноте переносится в основном ионами натрия, следующими по электрохимическому градиенту через катионные каналы наружного сегмента. Гиперполяризация фоторецептора при действии света обусловлена закрытием этих каналов, что приводит к сдвигу мембранного потенциала в сторону ЕК.
Свойства каналов фоторецептора
Катионные каналы наружного сегмента при физиологических условиях имеют соотношение в проводимости ионов кальция/натрия/калия как 12,5: 1,0: 0,7 и проводимость одиночного канала около 0,1 пСм21). Из-за того, что концентрация натрия гораздо выше, чем концентрация кальция, около 85% входящего тока приходится на ионы натрия. Движущая сила ионов калия направлена наружу клетки. При прохождении канала ионами кальция, эти ионы прочно связываются со стенкой поры и мешают, таким образом, проведению других ионов. Именно из-за этого удаление кальция из внеклеточной среды приводит к более легкому прохождению ионов калия и натрия через каналы, проводимость которых возрастает до 25 пСм.
Фесенко, Яу, Бейлор, Страйер с коллегами показали, что циклическая ГМФ играет роль внутреннего переносчика сигнала от диска к поверхности мембраны. Как показано на рис.4, высокая концентрация цГМФ в цитоплазме поддерживает катионные каналы в открытом состоянии. При уменьшении концентрации цГМФ с внутренней поверхности мембраны открытие катионных каналов становится редким событием. Таким образом, мембранный потенциал фоторецепторов является отражением концентрации цГМФ в цитоплазме: чем выше концентрация цГМФ, тем больше деполяризация клетки. Концентрация же цГМФ, в свою очередь, зависит от интенсивности падающего света. Повышение интенсивности света приводит к уменьшению концентрации цГМФ и уменьшает долю открытых каналов. В отсутствии цГМФ почти все каналы закрыты, и сопротивление мембраны наружного сегмента приближается к значению сопротивления липидного бислоя.
Молекулярная структура цГМФ-управляемых каналов
Была выделена кДНК для ионных каналов наружного сегмента палочек и определены аминокислотные последовательности субъединиц данных каналов в сетчатке человека, быка, мыши и курицы. Было обнаружено значительное сходство ДНК для этих и других цГМФ-управляемых ионных каналов - например, найденных в обонятельной
Передача сигнала в фоторецепторах
Каким образом фотоизомеризация родопсина приводит к изменению мембранного потенциала? На протяжении многих лет было понятно, что необходим какой-то внутренний посредник для возбуждения электрических сигналов в палочках и колбочках. Одной из причин подозревать, что информация о поглощении фотонов в области наружного сегмента палочки передается при помощи переносчика, был тот факт, что сам пигмент родопсин находится внутри диска, а сигнал распространяется через цитоплазму до внешней мембраны. Второй причиной было значительное усиление ответа. Бейлор с коллегами, изучая фоторецепторы черепахи, показал, что уменьшение мембранной проводимости и регистрируемых электрических сигналов возникает уже при поглощении всего одного фотона и активации одной из 108 пигментных молекул.
Последовательность событий, при которых активировавшаяся молекула фотопигмента изменяет мембранный потенциал, была выяснена при изучении наружных сегментов палочек и колбочек, используя методы двухэлектродной фиксации потенциала и методов молекулярной биологии. Схема передачи сигнала от поглощения фотона света до электрического сигнала показана на рис. 3.
В темноте в наружных сегментах палочек и колбочек протекает постоянный входящий "темновой" ток. В результате их мембран системе. Мембранные участки этих каналов проявляют структурные сходства с другими катион-селективными каналами, особенно в области S4 и в области, формирующей ионную пору. Ионные каналы фоторецептора являются тетрамерами, составленными, по крайней мере, из 2 различных белковых субъединиц с молекулярной массой 63 и 240 кДа соответственно.
Внутриклеточные места связывания нуклеотидов расположены около карбоксильного конца субъединиц. Экспрессия этих субъединиц в ооцитах приводит к формированию катионных каналов, имеющих свойства подобные каналам, расположенным в наружных сегментах палочек: они активируются цГМФ и имеют ожидаемые соотношения проводимости и проницаемости.
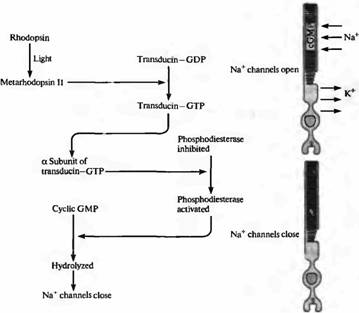
Рис. 6. Механизм активации G-белка при активации молекулы фотопигмента. G-белок трансдуцин связывает ГТФ в присутствии метародопсина II, что приводит к активации фосфодиэстеразы, которая, в свою очередь, гидролизует цГМФ. При падении концентрации цГМФ закрываются натриевые каналы.
Метаболический каскад циклического ГМФ
Цепочка событий, приводящих к уменьшению концентрации цГМФ и к последующему закрытию ионных каналов, показана на рис.5. Уменьшение внутриклеточной концентрации цГМФ вызывается светом, приводящим к образованию метародопсина II, промежуточного продукта в распаде фотопигмента. Метародопсин II воздействует, в свою очередь, на G-белок трансдуцин, который состоит из 3 полипептидных цепей·
Взаимодействие метародопсина II и трансдуцина приводит к замене связанной с G-белком молекулы ГДФ на ГТФ. Это активирует ?субъединицу G-белка, которая отделяется от субъединиц и, в свою очередь, активирует расположенную в примембранной области фосфодиэстеразу: фермент, который гидролизует цГМФ. Концентрация цГМФ падает, становится меньше открытых ионных каналов, палочка гиперполяризуется. Каскад прерывается при фосфорилировании С-конца активного метародопсина II. Ключевая роль цГМФ в регулировании состоянии катионных каналов была подтверждена биохимическими экспериментами. Освещение фоторецепторов может вызвать 20% падение уровня цГМФ в клетке.
Рецепторы позвоночных, деполяризующиеся при действии света
Интересным исключением из механизма фоторецепции, приведенного выше, являются некоторые рецепторы позвоночных. У ящериц есть третий глаз, расположенный на макушке. В нем находятся маленькие "колбочки", способные воспринимать изображение, сходное с воспринимаемым основными (боковыми) глазами. Эти фоторецепторы, однако, примечательны тем, что деполяризуются при освещении. Каналы, управляемые нуклеотидами, здесь имеют сходное строение и функционирование с фоторецепторами других позвоночных, за одним исключением: активация фоторецептора и G-белка приводит к увеличению концентрации цГМФ. В результате этого каналы наружного сегмента открываются, и катионы направляются внутрь клетки, формируя "световой" ток. Это происходит благодаря угнетению активности фосфодиэстеразы в темноте. В результате в глазу ящерицы происходит следующая последовательность событий: свет - [цГМФ] - ткрытие катионных каналов наружного сегмента - деполяризация.
Усиление сигнала в каскаде цГМФ
Двухэтапный каскад цГМФ обеспечивает значительное усиление исходного сигнала, чем и объясняется исключительная чувствительность палочек к свету. Во-первых, одна молекула активного метародопсина II катализирует присоединение многих молекул ГТФ вместо ГДФ и, таким образом, освобождает сотни субъединиц G-белка. Во-вторых, каждая субъединица активирует одну молекулу фосфодиэстеразы в диске, которая способна расщепить огромное количество молекул цГМФ, находящихся в цитоплазме, и таким образом привести к закрытию большого количества каналов.
Сигналы в ответ на одиночные кванты света
Данные о том, что одиночные кванты света могут вызывать воспринимаемое ощущение света, вызвали большое количество вопросов. Насколько велик этот единичный ответ? Каким образом этот сигнал выделяется из уровня шума? И каким образом такая информация достоверно передается из сетчатки в высшие зрительные центры? Чтобы измерить сигналы в ответ на одиночные кванты света, Бейлор с коллегами регистрировали токи от отдельных палочек в сетчатках жабы, обезьяны и человека. Эти опыты представляют собой уникальный пример эксперимента, в котором показано, как такой сложный процесс, как восприятие слабых вспышек света, может коррелировать с изменениями, происходящими на уровне отдельных молекул.
Процедура по изоляции фрагмента сетчатки от животного либо трупного материала должна для этих экспериментов производиться в темноте. Для измерения тока наружный сегмент палочки засасывается в тонкую пипетку. Как и ожидалось, эти эксперименты показывают, что в темноте ток постоянно течет внутрь наружного сегмента. Вспышки света приводят к закрытию каналов в наружном сегменте, вызывая уменьшение "темнового" тока. Амплитуда токов невелика и пропорциональна числу поглощенных квантов. Иногда вспышка вызывает одиночный ответ, иногда - двойной, а иногда - вообще никакой реакции.
В палочках обезьяны уменьшение тока в ответ на поглощение одного фотона, составляет около 0,5 пА. Это соответствует закрытию порядка 300 каналов, т.е. от 3 до 5% всех открытых в темноте каналов. Это достигается благодаря значительному усилению сигнала в метаболическом каскаде цГМФ. Более того, из-за крайней стабильности зрительных пигментов, упомянутой ранее, случайная изомеризация и ложное закрытие каналов - события очень редкие. Это приводит к тому, что эффекты отдельных квантов света выделяются на фоне очень низкого постоянного шума. Было показано, что электрическая связь посредством щелевых контактов между фоторецепторами обеспечивает дополнительный сглаживающий эффект, который уменьшает фоновый шум и улучшает отношение сигнал/шум ответов палочек на одиночные кванты.
Литература
1. Finn, J. T., Grunwald, M. E, and Yau, K-W. 1996. Cyclic nucleotide-gated ion channels: An ex-tended family with diverse functions. Annu. Rev. Physiol.58: 395-426.
2. Nakanishi, S., Nakajima, Y., Masu, M., Ueda, Y, Nakahara, K., Watanabe, D., Yamaguchi, S., Kawabata, S., and Okada, M. 1998. Glutamate receptors: Brain function and signal trans-duction.
Перепечатка материалов без ссылки на наш сайт запрещена