Учебное пособие: Методические рекомендации к определению и выведению гемограммы у животных
Учебное пособие: Методические рекомендации к определению и выведению гемограммы у животных
Министерство сельского хозяйства Российской Федерации
Ульяновская государственная сельскохозяйственная академия
Кафедра физиологии сельскохозяйственных животных и зоологии
Методические рекомендации к определению и выведению гемограммы у животных
Ульяновск, 2005 г.
УДК
Н.А.Любин, Л.Б.Конова.
Методические рекомендации к определению и выведению гемограммы у сельскохозяйственных и лабораторных животных при патологиях.
Для студентов и аспирантов факультетов ветеринарной медицины и технологического. Ульяновск, ГСХА, 2005, с.
При подготовке настоящих рекомендаций был использован опыт работы…
РЕЦЕНЗЕНТ: В.А.Ермолаев, доктор ветеринарных наук, профессор
Рекомендовано к изданию
методической комиссией
факультета ветеринарной медицины
Протокол № от 2005 г.
Ульяновская государственная сельскохозяйственная академия, 2005 г
МЕТОДИКА ПРИГОТОВЛЕНИЯ МАЗКОВ КРОВИ ДЛЯ ИССЛЕДОВАНИЯ
А. ВЗЯТИЕ КРОВИ И ПОЛУЧЕНИЕ МАЗКА
Циркулирующая в сосудистой системе кровь представляет более или менее равномерную взвесь в плазме форменных элементов: эритроцитов, лейкоцитов и кровяных пластинок (у птиц, рептилий и амфибий — тромбоцитов). Абсолютное количество и соотношение отдельных групп клеток в крови различных видов животных неодинаковы. Наибольшие колебания дают эритроциты: от 3,5 млн. в 1 мм 3 крови кур до 14,4 млн. в том же объёме крови коз. Несколько меньше видовые колебания количества белых кровяных телец: в 1 мм 3 крови млекопитающих содержится от 5 до 15 тыс. лейкоцитов. Количество лейкоцитов в крови птиц значительно выше: у кур, например оно, доходит до 35 тыс., а у гусей до 38 тыс. в 1 мм 3. Наконец, содержание кровяных пластинок в 1 мм3 крови колеблется от 200 до 400 тыс.
В крови, взятой из различных участков сосудистой системы, находится далеко не одина- ковое количество кровяных телец, особенно, и это наиболее важно, лейкоцитов. Значительно колеблется при этом и соотношение отдельных форм белой крови (табл.1,2).
Таблица1
Количество лейкоцитов в различных участках кровяного русла у кроликов
| Название органа или сосуда, ткани | % | Название органа или сосуда, ткани | % |
| Паренхима печени | 83,9 | Лёгкие . | 74,4 |
| Паренхима селезенки | 385, 3 | Vena. Pulmon. | 43,9 |
| Почки | 103,8 | Мышцы сердца | 61,7 |
| Надпочечники | 73,8 | Костный мозг | 128,29 |
| Vena mesent. | 89,0 | Vena. femoralis | 62,4 |
| Art. mesent. | 84,4 | Аrt . femoralis | 69,9 |
| Vena cava caud. | 4.9 | Vena. renalis | 60,5 |
Если принять среднее количество лейкоцитов в 1 мм3 крови из ушной вены за 100%, то в крови из сосудов других органов содержится:
Поэтому важно брать кровь для анализа всегда из одного и того же сосуда или группы сосудов. У всех сельскохозяйственных млекопитающих таким местом являются вены уха, у кур — гребень, у уток и гусей — мякоть ступни ноги.
Таблица 2 Состав белой крови свиней
| Кровь | Базофилы | Эозино-филы | Нейтрофилы | Лимфо-циты | Моноциты | ||
| Юные |
Палочко ядерные. |
Сегменто ядерные. |
|||||
| Из вены уха | 0,5 | 2,5 | 1,0 | 5,5 | 31,5 | 55,5 | 3,5 |
| Из сердца | до 0,1 | 0,4 | 1,0 | 10,0 | 62,0 | 24,5 | 4,0 |
Количество лейкоцитов зависит и от физиологического состояния животного.
У многих сельскохозяйственных и лабораторных животных заметно выражены пищеварительный лейкоцитоз (особенно у собаки), менее у лошади, и колебания количества лейкоцитов и (особенно) эритроцитов при мышечной работе. При некоторых патологических состояниях имеет место ретенция белых кровяных телец в сосудах печени и надпочечника. Лейкоцитоз наблюдается также во второй половине беременности.
Место взятия крови тщательно выстригается и если нужно, промывается водой с мылом, затем спиртом (или спиртом с эфиром). Рекомендуется тщательно растирать место взятия крови ваткой со спиртом и эфиром, что вызывает местную гиперемию и вместе с тем помогает избежать случайных колебаний лейкоцитарной формулы, связанных с некоторым застоем крови в мелких венах уха. Особенно следует иметь в виду возможное избирательное скопление в них эозинофилов.
Хорошие мазки крови можно получить только на очень чистых, обезжиренных предметных стёклах, тщательно промытых сначала горячей водой с мылом, а затем, после высушивания, — спиртом с эфиром. Промывание в спирте с эфиром особенно важно для полного обезжиривания стёкол. Если используются уже бывшие в употреблении стёкла, то их необходимо предварительно прокипятить в воде с содой.
Прокол тканей для получения крови лучше всего производить иглой Франка, но можно употреблять и обыкновенную иглу или специальное перо для уколов.
Первую выступившую на поверхность каплю крови быстро и тщательно стирают с места укола, а из второй и последующих приготовляют мазки.
Приготовление мазка крови. Чистое предметное стекло держат, как показано на рисунке 1, между большим и средним пальцами левой руки. В правой руке, теми же или большим и указательным пальцами держат чистое покровное или тонкое шлифованное предметное стекло. По крайней мере, одно ребро таких стёкол должно быть уже ширины того предметного стекла, на котором приготовляют мазок. Обычно это достигается подбором или обламыванием углов шлифованного стекла.
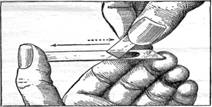
Рис. 1. Приготовление мазка крови.
Поверхностью предметного стекла, зажатого в левой руке, осторожно, но быстро касаются выступившей из прокола капли крови, стараясь сделать это ближе к среднему пальцу и, сейчас же, приведя стекло в горизонтальное положение, прикладывают к его поверхности узкое ребро того стекла, которое держат в правой руке. Приложенное ребро должно лежать перпендикулярно к длинным граням предметного отекла, а самое приложенное стекло нужно наклонить в сторону капли под углом в 40—50°. Держа это стекло, таким образом, осторожно двигают его в сторону капли до соприкосновения с нею. Как только капля, коснувшись подвижного стекла, разойдётся по линии соприкосновения стёкол, верхнее наклонное стекло быстрым, но ровным движением направляют обратно, в сторону большого пальца, сохраняя всё время прежний угол наклона в 40—50°. Полученный таким способом мазок высушивают на воздухе и на нём пишут иглой название или номер животного, его пол ((J или Q), и дату взятия крови.
Можно изготовлять мазки и на покровных стёклах. Для этого одним покровным стеклом берут очень маленькую каплю крови и прикладывают к нему другое покровное стекло так, чтобы углы одного стекла легли на середину рёбер другого. Как только капля крови разойдётся тончайшим слоем между обоими стёклами, последние параллельным
движением в противоположные стороны разводятся, и, таким образом, получаются два мазка.
В. ФИКСАЦИЯ МАЗКА
Чтобы закрепить все форменные элементы крови в препарате с максимально возможным сохранением их структуры и подготовить мазки к последующей окраске, существуют различные методы фиксации мазков. Наибольшее практическое значение имеют:
1. Фиксация абсолютным метиловым спиртом. Это лучший метод фиксации. Сухие мазки на 3 минуты погружаются в абсолютный метиловый спирт или на то же время спирт наливается на мазок, вполне покрывая препарат. Через 3 минуты мазки вынимают (или сливают с них спирт) и просушивают на воздухе.
2. Фиксация абсолютным этиловым спиртом, смешанным с равным количеством эфира. Фиксация длится 10—30 минут в обычных сосудах для гистологических растворов. Этот способ значительно хуже, так как даёт много артефактов.
В. ОКРАСКА МАЗКА. ОСНОВНОЙ МЕТОД ОКРАСКИ ПО РОМАНОВСКОМУ
Посредством окраски препарата наиболее отчетливо выявляется тончайшая структура, как ядра, так и цитоплазмы. Принцип современных методов окраски мазков крови открыт в 1891 г. Д. Л. Романовским и заключается в избирательном поглощении (химическом и коллоидальнохимическом) веществами клетки трёх красящих веществ — азура метиленовой синьки и эозина. Азур («красная из метиленовой синьки») имеет амфотерноосновную реакцию, метиленовая синька — щелочную, эозин — кислую.
Ядро клетки, богатое нуклеопротеидами и нуклеотидами, базофильно, т. е. окрашивается основными красками с избирательным поглощением а з у р а.
Цитоплазма молодых клеток, относительно богатая нуклеиновыми кислотами (нуклеотидами), также, хотя и в меньшей степени, базофильна. При этом избирательно поглощается преимущественно метиленовая синька. Цитоплазма же многих зрелых клеток крови (прежде всего нейтрофилов) ацидофильна (оксифильна).
Лимфоциты сохраняют базофилию цитоплазмы на всех стадиях развития, избирательно поглощая метиленовую синьку. Наличие базофилии молодой цитоплазмы, указывающее на относительное богатство её нуклеиновыми кислотами, связано с сохранением способности молодых клеток к интенсивному синтезу белков.
Лимфоциты сохраняют эту способность на весь онтогенез.
Базофилия гранул базофилов определяется наличием в них кислой слизи (мукоитиносерная кислота).
В лабораторной практике чаще всего пользуются следующими способами окраски по методу Романовского.
Окраска раствором Гимза
Краска Гимза, применяемая при окраске по методу Романовского, представляет собой комбинацию метилен-азура (азур II) и эозина (В, жёлтого). Она состоит из азура II*—3,0, эозина В —0,8, химически чистого глицерина 250,0 и метилового спирта 250,0.
Эта краска обычно отпускается готовой. Очень многое в успехе окрашивания определяет безупречное качество раствора краски, а последнее в высшей степени зависит от реакции воды.
Для приготовления рабочего раствора краски употребляется дважды дистиллированная вода. Обычно она имеет рН=5,4, т. е. слишком кисла, и даёт слабую, плохую окраску. Поэтому такую воду нужно подщелочить, прибавив к 2—3 л воды несколько капель 1-процентного раствора соды. Лучшим рН воды следует считать 6,8—6,9 (до 7,1). При более щелочной реакции воды мазки получаются более синими (даже эритроциты).
Практически пригодность воды к окраске определяют по растворению в ней гематоксилина. В 10 см3 дистиллированной воды (лучше всего в часовом стёклышке) кладут пинцетом несколько крупинок гематоксилина. Не ранее как через 1 минуту и не позже 5 минут вода должна окраситься в ясный слабофиолетовый цвет. Более раннее и интенсивное окрашивание указывает на сильно щелочную реакцию, более позднее — на кислую.
Если нет очень хорошего гематоксилина, то реакцию воды, подщелачиваемой раствором соды, можно определить, пользуясь в качестве индикатора нейтральной красной. В два химических стакана (или колбы Эрленмейера) наливают по 200 см3 дистиллированной воды и прибавляют 1 — 2 капли 1-процентного раствора нейтральной красной. При рН дистиллированной воды, равном 5,4—5,5, получается свекловично-красная (рубиновая) окраска. После этого в один из стаканов прибавляют по капле, тщательно размешивая, 1% раствор соды до появления слабого оранжевого оттенка (цвет лососины). Нельзя доводить воду до ясно оранжевого и, тем более, жёлтого цвета, так как в этом случае реакция будет сильно щелочной.
Более кропотливо пользоваться индикаторами Михаэлиса и менее точно — универсальным индикатором.
Лучше всего для получения нужной и стойкой реакции воды применять буферные растворы. Хорошими буферными растворами для этой цели будут фосфатные. Для них нужно иметь два исходных раствора:
1. Двуосновного фосфата натрия (Na2HP04. •2Н20)—17,814 г на 1 л (рН =8,302).
2. Одноосновного фосфата калия (КН2Р04) — 13,638 г на 1 л (рН =4,529).
Азур II — смесь в равных частях красок: азур I (диметилтиониохлорид) и метиленовой синьки. В постаревшем растворе исходной краски Гимза метиленовая синька часто плохо окрашивает цитоплазму лимфоцитов (нет чисто голубого цвета). Тогда необходимо добавить метиленовой синьки в исходный раствор краски.
Если к литру дистиллированной воды прибавить по 5 см3 того и другого раствора, то рН такой воды будет равен 6,813. Случайные колебания концентрации углекислоты в воздухе при таком способе не оказывают влияния на рН воды.
Перед окраской (ex tempore) к дистиллированной воде прибавляют исходный раствор краски Гимза из расчёта 1,0—1,5 капли краски на 1 см3 воды.
Рабочий раствор краски наливается пипеткой (осторожно!) на предметное стекло так, чтобы препарат (мазок крови) был полностью покрыт краской. Стёкла помещаются над широкой чашкой Петри на стеклянных палочках, соединённых резиновыми трубками попарно. Во избежание осадков, которые невозможно отмыть, можно стёкла укладывать мазками вниз, на стеклянные палочки, лежащие в чашке Петри, так, чтобы краска покрывала мазки полностью (способ академика Н. Д. Стражеско).
При первом способе на один мазок тратится 2,5— 3,0 см3 рабочего раствора краски.
Через 20—30 минут краску сливают, препараты промывают водопроводной чистой водой и высушивают на воздухе. Для окраски старой краской требуется меньше времени.
При окраске раствором Гимза хорошо дифференцируется структура ядра, несколько хуже — структура цитоплазмы, особенно нейтрофильная зернистость. Однако при очень умелом окрашивании и зернистость выявляется достаточно хорошо.
По этому методу плохо окрашиваются псевдоэозинофилы (палочкозернистые гранулоциты) крови сельскохозяйственных птиц.
Окрашенные краской Гимза ядра имеют красивый фиолетово-красный цвет (цвет вишни); нейтрофильная зернистость — розовато-фиолетовый; эозинофильная зернистость — розовый или красно-розовый; базофильная —цвета мальвы; азурофильная — красно-фиолетового цвета.
Цитоплазма лимфоцитов — голубая, моноцитов — от голубовато-серой до пепельно-серой.
Эритроциты — красновато-розового цвета, полихроматофилъные эритроциты — синеватые, базофильная пунктация эритроцитов — синяя.
Если нет хорошей готовой краски Гимза, то можно хорошо окрасить мазки по Н о х т у. Для окраски по Нохту приготовляется два раствора: 1) 1-промил-льный раствор азура II; 2) 1-промилльный раствор эозина. Перед приготовлением рабочего раствора краску нужно оттитровать. Для этого сначала берут на 3 см3 дистиллированной воды 4 капли раствора азура II и 3 капли раствора эозина (т. е. в отношении 4 : 3). Если окраска мазков неправильна, пробуют отношение 4 : 4, или 4 : 5, или 4 : 6, и т. д.
Достаточно интенсивное окрашивание получается, если на 1,5 см3 азура II берётся 1,0 см3 эозина и 6 см3 воды.
Модификация Паппенгейма (Май-Грюнвальд — Гимза)
При этом способе окраски предварительная фиксация мазка не нужна, так как первая краска — Май-Грюнвальд имеет растворителем метиловый спирт.
Метод окраски двухмоментный. Сперва мазок покрывают 2 см3 неразведённой краски
Май-Грюнвальд, представляющей собой раствор в метиловом спирте эозина и метиленовой синьки. Если нет готовой надёжной краски, то растворяют приготовленный фабричным способом сухой порошок Май-Грюнвальд: 1,0 г порошка на 100 см3 абсолютного метилового спирта и 50 см3 чистого глицерина. Хорошая краска получается и без глицерина.
Через 3 минуты к краске Май-Грюнвальд, находящейся на мазке, прибавляют 2 см3 дистиллированной воды и тщательно смешивают их продуванием или последовательным набиранием и выпусканием через тонко оттянутую пипетку. Когда, примерно через 1 минуту, мазок приобретает розовый оттенок, краску с препарата сливают и после этого, не высушивая, 10—12—15 минут красят мазок рабочим раствором краски Гимза, а затем промывают чистой водопроводной водой. Этот способ окраски удачно сочетает хорошее выявление зернистости клеток краской Май-Грюнвальд с чёткой окраской структуры ядра раствором Гимза.
Такая окраска особенно ценна для выявления псевдоэозинофилов (специальных гранулоцитов) куриной крови.
Результаты окраски: ядра красно-фиолетовые, цитоплазма лимфоцитов голубовато-синяя, азурофильная зернистость лимфоцитов пурпурно-красная, миелоидная азурофилия фиолетово-коричневая, так же как и центральная субстанция кровяных пластинок. Нейтрофильная зернистость коричневато-красная, до синевато-розовой; эозинофильная — от красно-оранжевой до кирпично-красного цвета; базофильная зернистость тёмного ультрамариново-фиолетового цвета (метахромазия); эритроциты медно-розовые; полихроматофильные эритроциты синеватые; базофильная пунктация эритроцитов (слабо выявляемая) синего цвета *.
В целом, мазки, окрашенные по этому способу, несколько богаче оттенками, чем при окраске раствором Гимза, но кажутся более тёмными, «мрачными».
Цветные таблицы картин крови выполнены с препаратов крови, окрашенных по Романовскому в модификации Паппенгейма, лучше всего диференцирующей структуру клеток.
Ускоренная окраска раствором Гимза (новая модификация)
Этот метод даёт хорошие результаты только в руках опытного исследователя. Преимущество — быстрота приготовления препарата, так как фиксация и окрашивание происходят одновременно.
Для работы необходимы капельницы ёмкостью в 30 см3, в которые наливается исходный раствор краски Гимза, разведённый пополам метиловым спиртом или чистейшим ацетоном. В хорошо закупоренной склянке этот раствор может сохраняться месяцами.
Окрашивание производится или в чашках со специальными перекладинками, или в обычных чашках Петри. На свежий мазок (не позднее 2-дневного!), высушенный на воздухе и не фиксированный, наливают из капельницы около 20 капель краски. Чтобы предохранить краску от испарения, чашка, в которой лежат препараты, закрывается стеклом.
Через 1/2 минуты — 1 минуту к краске на мазке приливают около 10 см3 подщелочённой дистиллированной воды (1—2 капли 1-процентного раствора щелочи на 50 см3 воды). Покачиванием чашечки хорошо перемешивают краску с подлитой водой. Через 10— 15 минут воду с краской сливают и мазок промывают водопроводной (недистиллированной!) водой.
В случае приготовления основного раствора с ацетоном особенно хорошо выявляется зернистость структура ядер кровяные пластинки.
• Описание цветов дано, с некоторыми небольшими изменением, по А. II. Крюкову.
Окраска крови и кровепаразитов по Г. Эпштейну
Фиксация метиловым спиртом
Готовят два раствора.
1-й: дистиллированной воды 100 см3; лимоннокислого лития — 1 г; толуидиновой голубой — 1г. После разведения профильтровать.
2-й: насыщенный водный раствор пикриновой кислоты.
Мазки красят 20—30 минут в первом растворе и, ополоснув в проточной воде, помещают на 1—2 секунды во второй раствор (пикриновой кислоты). Промывают (тщательно!) снова в водопроводной воде и после этого быстро обсушивают фильтровальной бумагой.
Окраска: эритроциты зелёные; ядра лейкоцитов фиолетово-синие; базофильная зернистость вишнёвая; эозинофильная зернистость изумрудно-зелёная; нейтрофильная зернистость серая; у кровепаразитов ядро красное, а цитоплазма синяя.
Окраска составом Лейшмана
На нефиксированный препарат налить 15—20 капель готового раствора краски Лейшмана (0,1 г порошка краски растворить в 10 см3 метилового спирта). Через 2,5—3 минуты прибавить 30 капель дестиллированной воды и красить ещё 5 минут. Промыть в течение 2—3 минут проточной водой и высушить на воздухе.
Специальные способы окраски и фиксации мазка
а) Получение и окраска толстой капли. На хорошо промытое (подщелочённой водой, затем спиртом с эфиром) предметное стекло наносят 2 крупные капли крови. Собственно, достаточно одной капли, но вторая служит «резервом» на случай неосторожного стирания одной капли, неудачи окраски и т. д. Каждая капля сейчас же, с помощью иглы, распределяется по стеклу ровным слоем — примерно в 1\4 - 1\2 мм толщины, чтобы получились пятна размером с десятикопеечную монету. Препараты тщательно высушиваются (в термостате при 37° или на солнце) в течение получаса.
После этого высохший препарат дважды окрашивают рабочим раствором Гимза (1 капля исходного спиртового раствора Гимза на 1 см3 дистиллированной воды).
Первое окрашивание длится около 3 минут после того, как в растворе краски появится красное облачко растворившегося гемоглобина. Затем один конец предметного стекла слегка приподнимают и старый раствор Гимза заменяют новым, приливая его очень осторожно с приподнятого конца препарата, в то время как прежний раствор краски с гемоглобином стекает с другого конца. Когда таким образом вся старая краска сменена новой, предметное стекло снова устанавливают горизонтально и продолжают докрашивание ещё 25 минут.
Значение метода состоит в том, что в толстой капле удается обнаружить таких паразитов крови, которые находятся в ней в небольших количествах. Быстро устанавливается наличие или отсутствие эозинофилов и производится их подсчет их подсчёт.
То же самое в отношении базофильных эритроцитов.
б) Оксидазная и пероксидазиая реакции лейкоцитов. Оксидазная реакция основана на возникновении индофеноловой голубой краски при воздействии окисляющих ферментов на анафтол и диметилпарафенилендиамин; в местах локализации оксидаз в клетке возникает синее окрашивание.
Техника реакции состоит в следующем:
1. Фиксация мазка в смеси из 1 части 40-процентного формальдегида и 9 частей 95-процентного спирта в продолжение нескольких секунд пли 40-процентного формальдегида и абсолютного алкоголя аа в продолжение 15—20 минут.
2. Окраска: а) 3 минуты слегка разведённым 1-процентным водным щелочным раствором анафтола (раствор приготовляется следующим образом: анафтол при нагревании в дестиллированной воде поднимается кверху и плавает в жидком виде; после введения в раствор кристалла едкой щёлочи анафтол растворяется в воде); б) не удаляя анафтола, на препарат наслаивают 1-процентный водный раствор диметилпарафенилендиамина. Через несколько минут зернистость лейкоцитов, содержащая оксидазу, становится темносиней. Докрашивается препарат сильно разведённым раствором фуксина Циля.
Пероксидазная реакция.
а) Окраска по Край-биш в модификации Грэма. Хорошо высохший мазок в течение 10—15 минут фиксировать жидкостью, состоящей из 1 части 40-процентного формалина и 9 частей 95-процентного спирта. После фиксации слегка промыть водой и покрыть раствором бензидина (приготовление: к 10 см3 40-процентного этилового спирта прибавляется несколько кристаллов бензидина +00,2 см3 3-процентной перекиси водорода). Окраска длится 5 минут. Затем смыть водой.
Места локализации пероксидаз окрашиваются сначала в сероватый, а затем в золотисто-коричневый цвет.
Последующее докрашивание — тионином, метиленовой синькой или краской Гимза.
б) Окраска по Сато. Воздушносухие мазки фиксируют в течение 30 секунд 1/2-процентным раствором медного купороса (CuS04) и затем окрашивают бензидином с перекисью водорода (рецепт приготовления — как в предыдущем методе). Через 2 минуты препарат осторожно промывают дистиллированной водой, на мазок наливают слой 1-процентного водного раствора сафранина, через 15—20 минут сафранин смывают водой и высушивают препарат на воздухе (избегать высушивания фильтровальной бумагой!).
Пероксидазо-положительные гранулы окрашиваются в темносиний цвет, ядра — в красно-жёлтый; эритроциты не окрашиваются.
Посредством оксидазной и пероксидазной реакций облегчается диференциация миэлоидных клеток от лимфоидных. Обе реакции дают аналогичные результаты, но оксидазная реакция более чувствительна.
Нейтрофилы и эозинофилы реагируют резко положительно, базофилы так же, но только на ранних стадиях развития. Зрелые формы оксидазо-отрицательны. Однако ряд авторов считает, что и зрелые базофилы оксидазо-положительны.
Лимфоциты дают безусловно отрицательную реакцию. Моноциты — иногда очень слабо положительную.
С клинической точки зрения представляет интерес тот факт, что при некоторых инфекционных заболеваниях у нейтрофилов исчезают положительная оксидазная п пероксидазная реакции.
в) Окраска токсической зернистости раствором Гимза при рН =5,4. Токсическую зернистость, в отличие от обычной, нормальной зернистости гетерофилов (нейтрофилов), избирательно окрашивают, при окраске по принципу Романовского раствором краски Гимза, применяя буферный раствор с рН =5,4.
Буферный раствор:
Едкий натр (химически чистый) . 21,6 г
Уксусная кислота (химически чистая) .............. 27,0 »
Дистиллированная вода до.....1000,0 см3
Приготовление краски:
Исходной краски Гимза …………10см3
Дистиллигрованной воды ……….40 »
Буферного раствора до ………….100 »
Свежие мазки окрашивают в продолжение 1 часа, старые препараты — дольше (до 2 часов). Краска с мазка смывается буферным раствором и затем высушивается, как обычно.
При окрашивании препарат нужно класть на раствор краски мазком вниз.
Белые кровяные тельца – лейкоциты.
Лейкоциты (белые кровяные тельца) различаются между собой как морфологически, так и по биологической роли в организме. Будучи полноценными клетками, имеющими протоплазму и ядро, лейкоциты обладают отчётливо выраженной способностью к активному способу питания путём захвата и внутриклеточного переваривания попадающих в кровь органических тел. Эта способность приобретает первостепенное биологическое значение в случае проникновения в организм патогенных микробов: пожирание их лейкоцитами — фагоцитоз (Мечников, 1882—1893) — составляет важнейшее средство борьбы организма с инфекцией.
Наряду с фагоцитозом, весьма важное значение имеет образование лейкоцитами иммунных тел. У многих низших, а весьма возможно и высших животных особые лейкоциты выполняют также функцию переноса питательных веществ (трефоциты). Наконец, отдельные виды лейкоцитов (эозинофилы высших животных) способны обезвреживать токсины. Крупную роль лейкоциты играют в обмене веществ и в образовании так называемых трефонов — стимуляторов клеточного роста, особенно в условиях регенерации тканей.
Структурные различия отдельных видов белых кровяных телец изучены, начиная с работ П. Эрлиха (Р. Ehrlich, 1877—1898 гг.), достаточно хорошо. Значительно менее изучены их функциональные особенности, их целлюлярная физиология. Несмотря на огромное количество работ, онтогенез белой крови полностью ещё не выяснен. Наконец, сложная нейро-гуморальная регуляция сосудистой и внесосудистой белой крови исследована в чрезвычайно малой степени. Мало данных имеется даже о длительности жизни белых кровяных телец. По некоторым авторам, она весьма невелика (3—4 дня).
Основным принципом современной классификации лейкоцитов является морфологический.
У различных сельскохозяйственных и лабораторных животных один и тот же тип лейкоцитов (особенно эозинофилы и нейтрофилы, или гетерофилы) имеет специфические отличия в структуре. Однако в главном структура каждого типа лейкоцитов у всех сельскохозяйственных животных весьма близка и поэтому целесообразно вначале дать их общее описание, без видовой дифференциации.
По структуре ядро эозинофилов близко к ядру нейтрофилов, но несколько бледнее и выглядит грубее, так как чередующиеся светлые (оксихроматин) и тёмные (базихроматин) участки ядра эозинофилов крупнее, чем у нейтрофилов.
По мере созревания клетки ядро эозинофильных лейкоцитов изменяется в том же направлении, что и нейтрофильных, т. е. ядерный жгут скручивается и утончается, сперва равномерно (юные и палочкоядерные формы),а затем отдельные участки (сегменты) почти перестают утончаться и остаются сравнительно толстыми, а находящиеся между ними — превращаются в "тончайшие нити (сегментоядерные формы). Однако сегментация ядра эозинофилов выражена не очень резко. Очень частой, типичной формой является 2-дольчатая форма ядра, причём дольки напоминают формирующиеся и только что отрывающиеся капли, обращенные друг к другу узкими концами, соединёнными перемычкой, или две груши, соединённые плодоножками. У овец полиморфность ядра эозинофилов выражена сильнее. Хотя при некоторых болезнях можно наблюдать в крови изменение отношения между возрастными формами эозинофилов в сторону увеличения более молодых (палочкоядерных, юных и даже миэлоцитов. — «сдвиг ядра влево»), но, ввиду относительной малочисленности (3—10%) эозинофилов, учёт ядерного сдвига в лейкоцитарной формуле не производится.
Эозинофилы имеют очень большое клиническое значение. Эозинофилы или исчезают из крови (анэо-винофилия), или уменьшаются в количестве (гипоэозинофилия), или, наконец, количество их резко нарастает (гиперэозинофилия, или просто эозинофилия). Большинство инфекционных заболеваний в первом своём периоде связано с резким уменьшением количества эозинофилов (гипоэозинофилия). Возврат эозинофилов в кровяное русло считают признаком ослабления болезни. При роже свиней и при многих инвазиях (особенно гельминтозах) наблюдается резкое увеличение эозинофилов (эозинофилия), доходящее у крупного рогатого скота до 40%. Эозинофилия встречается и при аллергических реакциях, причём здесь её связывают, так же как и при гельминтозах, с раздражением системы блуждающего нерва.
Функции эозинофилов недостаточно изучены.
Вероятна способность их зернистости к обезвреживанию токсинов, а также участие зёрен в окислительных процессах. Эозинофилы скопляются в местах тканевой регенерации. Характерна локальная эовинофилия кишечника.
III. Специальные зернистые лейкоциты, или нейтрофилы
Нейтрофилы (специальные зернистые лейкоциты, гетерофилы, псевдоэозинофилы или амфиоксифилы некоторых животных) имеют очень важное значение для клиники и физиологии.
Клетки нейтрофилов округлые, диаметром от 7,0 до 15,0 µ. В цитоплазме обильная, очень мелкая, нейтрофильная зернистость. Я д р о, по мере развития клетки, постепенно сегментируется. Зернистость ясно заметна даже в свежей неокрашенной крови. Эти при жизни клетки серебристо блестящие зёрнышки густо выполняют эндоплазму, передвигаясь с нею при амебоидных движениях клетки. Эктоплазма представляет собой тонкий гомогенный периферический слой, свободный от гранул. У некоторых животных (мыши, крысы и кошки) эерни-стость выражена очень слабо, но, однако, вопреки отрицанию Максимова, несомненно есть.
Цитоплазма оксифильна, окрашивается в бледнорозовый цвет, иногда почти бесцветна. Изредка в цитоплазме нейтрофилов встречаются небольшие участки, сохранившие базофилию, характерную для материнской клетки. Такие ясноголубые пятна получили название телец Деле (Dohle).
Окраска зернистости специальных гранулоцитов несколько различна у разных видов животных. Поэтому в последнее время для нейтрофилов предложено новое, удачное наименование — гетерофилы.
У обезьяны, собаки, кошки и свиньи зернистость имеет сродство к нейтральным краскам и при комбинациях красок по Романовскому окрашивается в розово-фиолетовый цвет. У большинства же остальных млекопитающих зернистость амфофильна, т. е. красится как кислыми, так и основными красками. У коровы, овцы, лошади и морской свинки (по Максимову) эти зёрна амфооксифильны, т. е. имеют большее сродство к кислым краскам; у кролика они окрашиваются эозином в яркокрасный цвет и поэтому называются псевдоэозинофилами.
У некоторых животных гранулы амфобазофильны. Зернистость псевдоэозинофилов кур и других домашних птиц окрашивается в яркокрасный цвет,весьма крупна и в большинстве случаев (особенно у зрелых форм) имеет палочковидную и даже веретенообразную, с заострёнными концами, форму. У более молодых форм зёрна округлы (Лебедев). Ряд авторов (Букраба, Я. Соловей) считает, что у кур нет вообще псевдоэозинофилов, а только эозинофилы. Но большинство исследователей диференцирует эозинофилов от псевдоэозинофилов, и морфологически и функционально, сближая последних со специальными гранулоцитами (Максимов, Рухлядев, Клинебергер и Карл, Лебедев). Особенно тщательное исследование разницы между эозинофильной и псевдоэозинофильной зернистостью произвёл Лебедев (1940 г.). Он установил, что при суправитальной окраске бриллианткрезиловая голубая окрашивает гранулы эозинофилов в голубовато-розовый цвет, а псевдоэозинофилов — в зеленовато-синий. Оксидазо- и пероксидазо-положительными оказались лишь эозинофилы. Окраска на липоиды по Зерту (Sehrt) дала положительный результат только с зернистостью эозинофилов. При обработке окрашенных препаратов смесью уксусной кислоты и спирта, зёрна эозинофилов сохраняют свою окраску, а верна псевдоэозинофилов обесцвечиваются. Наконец, сдвиг ядра псевдоэозинофилов у кур был идентичен с закономерностями сдвига ядра нейтрофилов при ряде патологических состояний у других животных. Следует, однако, отметить, что ранние стадии развития псевдоэозинофилов и эозинофилов, окрашенные растворами Гимза или по Паппенгейму, различать крайне трудно.
Разделение лейкоцитов на типы можно представить так:
| Лейкоциты (белые кровяные тельца) | ||||
| Гранулоциты | Агранулоциты | |||
| Имеют цитоплазматическую зернистость. Содержат оксидазу. По типу окислительного обмена отличаются более интенсивным поглощением кислорода и значительным анаэробным гликолизом | Не имеют цитоплазматической зернистости или имеют мелкую азурофильную зернистость, не определяющую функциональную значимость клетки. Оксидазы не содержат или содержат только следы по типу окислительного обмена менее интенсивно поглощают кислород и обладают вдвое слабейшей способностью к анаэробному гликолизу. | |||
|
Базофилы или тучные клетки (с базофильностью в цитоплазме) |
Эозинофилы (с ацидофильной зернистостью в цитоплазме |
Нейтрофилы (гетерофилы), или специальные гранулоциты (с нейтрофильной зернистостью в цитоплазме) | Лимфоциты (цитоплазма голубая с перинуклеарной зоной. Округлое , темно прокрашивающееся ядро, относительно грубой структуры) | Моноциты (дымчато-серая, иногда с розоватым или лиловатым оттенком цитоплазма, несколько расчлененное, бледно окрашивающееся, тонкой структуры ядро) |
Следует иметь в виду, что так называемая «структура» ядер является, при обычных способах фиксации, в том числе и фиксации метиловым спиртом, в значительной степени результатом коллоидальной флокуляции веществ ядра. В зависимости от применяемых фиксаторов, эта структура существенно изменяется. На самом же деле, как показал П. В. Макаров (1948 г.), покоящееся нативное ядро — за исключением ядрышка — оптически пусто, без микроструктур. В период кариокинетического деления в нём возникают временные образования — хромосомы.
Поэтому в дальнейшем описании под структурой ядра следует понимать возникающие в ядре при взаимодействии с фиксирующими и красящими веществами варьирующие образования — коагуляты. В ядрах различных клеток, в зависимости от специфических коллоидально химических различий их ядерной плазмы (кариоплазмы), эти коагуляты имеют некоторые морфологические особенности, которые и дают возможность различать между собою виды клеток.
А. ГРАНУЛОЦИТЫ
I. Базофилы
Базофильные гранулоциты, или тучные клетки, обычно круглой или округло-овальной формы, диаметром 8—15 µ. (у лошадей и коров несколько более крупные). Сама цитоплазма слабооксифильна и окрашивается в бледный, розовато-фиолетовый или, иногда. сыровато-голубой цвет, но находящиеся в ней крупные округлые зёрна (гранулы) резко базофилъной природы и окрашиваются метахроматически в тёмный красно-фиолетовый или ультрамариново-фиолетовый цвет (цвет мальвы — по Крюкову).. Зёрна легко растворяются в воде и потому в препаратах, фиксированных плохо обезвоженным метиловым спиртом, часто на место верен в цитоплазме образуются белые «окошечки». При фиксации абсолютным метиловым спиртом зёрна сохраняются хорошо. Расположение гранул в цитоплазме неравномерное, рыхлое. Часто они закрывают отдельные участки ядра. По своей химической природе базофильные зёрна являются белками, близкими к гликопротеидам.
Ядро базофилов — неясной структуры, неправильно лопастное или округлое, окрашивается в фиолетово-розовый цвет. В ядре расплывчато чередуются более светлые поля оксихроматина с темноокрашенными базихроматиновыми полями.
В базофилах чрезвычайно трудно различить стадии миэлоцита — юную, палочкоядерную и сегментоядерную. Вообще сегментированность ядра выражена слабо. Практического значения, для подсчёта лейкоцитарной формулы, дифференциация базофилов по степени их зрелости не имеет, прежде всего, потому, что в крови млекопитающих их очень мало: от 0,1 до 1—2%, в среднем 0,5%. Кровь сельскохозяйственных птиц содержит 3—4% базофилов, а содержание их в крови лягушек доходит до 23%.
Вообще, содержание базофилов очень высоко у амфибий, рептилий и у некоторых рыб.
Функциональное значение базофилов не выяснено. Повидимому, они играют некоторую роль в защите организма при парентеральном введении чуждых белков. Они способны фагоцитировать и содержать окислительные ферменты. Ряд учёных считает их трефоцитами («питающие клетки» Либмана). Такие клетки, переносящие питательные вещества, особенно широко распространены у беспозвоночных, где они часто преобладают.
Клиническое значение базофилов невелико. Количество их несколько возрастает при инъекции белков, при некоторых авитаминозах (группы В) и гепатических циррозах.
II. Эозинофилы
Эозинофилы (синонимы — оксифилы или ацидофилы) — это крупные (особенно у лошади) круглые клетки, диаметром от 8,2 до 19,8)1. Очень редко попадаются карликовые формы эозинофилов (особенно у крупного рогатого скота при депрессии гемопоэза). Очень крупны Эозинофилы лошади.
Цитоплазма слегка базофилъна, бесцветна или голубоватого цвета. Зёрна ярко окрашены эозином в интенсивный красный или розово-красный цвет (по описанию Крюкова, в кирпично-красный). У птиц они скорее розовые, чем красные. У молодых форм гранулы часто окрашены базофильно и лишь постепенно, по мере созревания клетки, становятся ацидофильными. У кошек цвет верен красновато-пурпурный.
Размерь и форма гранул весьма различны. У лошади .они очень крупные (до Зµ в поперечнике], покрывают часть ядра и придают эозинофилу вид плода малины (табл. 1—2). Довольно крупные верна у эозинофилов кролика (до 1,5 µ). Значительно мельче эозинофильная зернистость овцы. У свиньи зерна очень правильной круглой формы.
Обычно зёрна эозпнофилов расположены очень тесно, у лошади они часто даже сдавливают друг друга и приобретают угловатую форму и между зёрнами трудно различить цитоплазму. Однако у некоторых животных (например, у овцы) зерна могут быть расположены сравнительно редко, особенно в молодых клетках, и тогда цитоплазма видна хорошо. Типичные эозинофилы имеются в крови почти у всех позвоночных (кроме некоторых рыб). У п т и ц (Казаринов, Лебедев) зёрна эозинофилов относительно мелки. У рептилий эозинофилы составляют большинство лейкоцитов. Ацидофильные зёрна эозинофилов рептилий, плотно расположенные в цитоплазме, то шарообразны, то овальны, иногда имеют форму ромбических кристаллоидов или, наконец, представляют собой глыбки неправильной формы. Эозинофилы амфибий весьма напоминают собой аналогичные клетки у млекопитающих. Их гранулы относительно весьма велики. У большинства рыб имеются типичные эозинофилы, чаще всего с простым, круглым ядром. У некоторых видов рыб эозинофилы атипичны, — это лимфоидные клетки с редкими, но очень крупными гранулами, в цитоплазме. Наконец, в крови некоторых видов рыб эозинофилы, невидимому, не содержатся.
Микрохимическими методами установлена липоидно-белковая природа зёрен эозинофилов; они содержат фосфор и, возможно, железо. По Кальману (Kallman), юные эозинофилы птиц имеют гранулы нуклеопротеидной природы; позднее они становятся чисто альбуминовыми.
Зёрна эозинофилов видны даже в неокрашенных клетках, где они выделяются жёлтым цветом и высоким показателем преломления.
У амфибий, особенно лягушек, при хорошей фиксации и окраске по Романовскому в модификации Паппенгейма, зернистость удаётся выявить достаточно ясно. Зернистость гетерофилов у рептилий выражена слабо, но утверждение Максимова, что цитоплазма гетерофилов амфибий красится диффузно или выявляет сетчатое строение, но не содержит различных гранул, несомненно неправильно.
Окрашиваемость гранул в гетерофилах рыб сильно варьирует; у одних видов зернистость нейтрофильна, у других амфофильна. Наконец, у некоторых видов рыб гетерофилы (типичные по сегментированному ядру) до сих пор не найдены.
Форма ядра специальных гранулоцитов изменяется в зависимости от
возраста клетки. Редко появляющаяся в крови (только при патологических состояниях)
начальная форма — миэлоцит имеет округлое, реже с отдельными вдавленнями, ядро.
В дальнейшем оно вытягивается («скручивается», по А. Н. Крюкову) в сочное
бобовидное или колбасовидное ядро (юная форма), а  затем ещё более
вытягивается и изгибается то в форме изогнутой палочки, то подковы или буквы S. Это палочкоядерная
форма. Наконец, ядро перекручивается и образует ряд сегментов (долек),
связанных очень тонкими, иногда почти незаметными нитями. Это
полиморфноядерные или сегментоядерные формы. Так как первые исследователи не
замечали перетяжек между сегментами и каждый сегмент принимали за отдельное
ядро (Эрлих), то эти клетки получили сначала название полинуклеаров
(многоядерных). В настоящее время их правильнее называют полиморфноядерными
нейтрофилами. Описанный процесс изменения ядра наблюдался у свиньи, собаки и
морской свинки.
затем ещё более
вытягивается и изгибается то в форме изогнутой палочки, то подковы или буквы S. Это палочкоядерная
форма. Наконец, ядро перекручивается и образует ряд сегментов (долек),
связанных очень тонкими, иногда почти незаметными нитями. Это
полиморфноядерные или сегментоядерные формы. Так как первые исследователи не
замечали перетяжек между сегментами и каждый сегмент принимали за отдельное
ядро (Эрлих), то эти клетки получили сначала название полинуклеаров
(многоядерных). В настоящее время их правильнее называют полиморфноядерными
нейтрофилами. Описанный процесс изменения ядра наблюдался у свиньи, собаки и
морской свинки.
У большинства сельскохозяйственных животных процесс созревания сопровождается не сегментацией ядра, а образованием колец и приводит к возникновению так называемых цепочкообразных и узловатых форм ядра. Это видно из следующего рисунка (рис. 2).
Ядро специальных гранулоцитов окрашивается интенсивно (особенно у молодых), с резким чередованием базихроматина и оксихроматина (тёмных и светлых участков). Поэтому у зрелых форм структура ядра грубая. В ядре относительно много базихроматина (нуклеопротеидов и нуклеиновых кислот).
Специальные гранулоциты — это микрофаги И. И. Мечникова. Он объясняет перешнуровывание и сегментацию их ядра как специальное приспособление к диапедезу (миграции с проникновением через стенки капилляров). Именно поэтому они получили название специальных гранулоцитов (А. Максимов).
Гетерофилы содержат оксидазу и протеолитические ферменты (трипсин). Но некоторым данным, содержание ферментов, особенно трипсина, увеличивается при преобладании в пище белков.
Количество специальных гранулоцитов в крови довольно велико и колеблется в зависимости от вида животного, его функционального состояния и заболевания. Больше всего их у собак (60—70% всех лейкоцитов), меньше всего — у крупного рогатого скота (25—35%).
При патологических состояниях организма состав специальных гранулоцитов значительно изменяется. Резко уменьшается количество сегментоядерных клеток и нарастает количество палочкоядерных, юных и даже миэлоцитов, мобилизуемых из костного мозга в сосудистую кровь. Так как в самой левой графе лейкоцитарной формулы отмечаются наиболее молодые, в нормальной крови не встречающиеся клетки — миэлопиты, а все более взрослые формы — юные, палочкоядерные и сегментоядерные — размешаются в соответствующих графах все правее, то обогащение крови более молодыми формами получило название «сдвига ядра влево».
Регенеративный и дегенеративный сдвиги ядра. Различают два основных типа сдвига ядра: а) регенеративный и б) дегенеративный.
а) Регенеративный сдвиг ядра выражается в сдвиге ядра влево с увеличением в крови палочкоядерных, юных и даже миэлоцитов; обычно при этом наблюдается лейкоцитоз. Этот сдвиг и усиление лейкопоэза являются показателем раздражения костного мозга, происходящего при его функциональной достаточности. Костный мозг, компенсируя гибель нейтрофилов в борьбе с инфекцией, отдаёт в кровяное русло, наряду со зрелыми, всё возрастающее количество недостаточно зрелых форм, обычно не поступающих в сосудистую кровь.
б) При дегенеративном сдвиге общее часто уменьшается, отмечается нарастание палочкоядерных форм, в значительной степени дегенеративных без дальнейшего сдвига ядра влево. Дегенеративный сдвиг является показателем функциональной недостаточности костного мозга, в котором наблюдается тканевая дегенерация.
Индексом сдвига ядра называется отношение (М+Ю+П)/С равное обычно для крови взрослой лошади (О+О+4)/50 = 4/50
Для крови верблюда он равен 12.5/38 , коровы 6/25 и свиньи 3/40 (по Сёмушкину и Домрачеву).
В легких случаях патологического процесса сдвиг ядра влево не идёт далее увеличения палочкоядерных и частично юных форм. Напротив, появление большого количества миэлоцитов и юных специальных гранулоцитов в крови свидетельствует о тяжести заболевания.
При некоторых заболеваниях (особенно кроветворных органов) в крови появляются гигантские полисегментированные клетки. У некоторых животных, однако (например, у овцы), полисегментированные нейтрофилы находятся и в нормальной крови.
К дегенеративным изменениям специальных гранулоцитов относятся: пикнотичность и причудливые, резко угловатые формы ядра, токсическая зернистость и многочисленные вакуоли в цитоплазме.
Особенно большое значение имеет токсическая зернистость цитоплазмы. При обычной окраске растворами Гимза или Паппенгейма, мелкая в физиологической норме зернистость резко укрупняется, и зёрна часто сливаются в причудливую сеть (токсически изменённая зернистость). Для удобства дифференциации нормальной зернистости от токсической лучше применять окрашивание карболфуксинметиленовой синькой по Е. Фрейфельд. В этом случае физиологически нормальная зернистость гетерофилов почти не окрашивается, а патологическая резко выступает в виде фиолетово-синих зёрен или нитей и сеток на нежнорозовом фоне цитоплазмы. Можно также применять окраску по Гимза при кислой реакции воды (рН=5,4).
Вакуоли довольно часты в токсически изменённых или «старых» гетерофилах.
Иногда при инфекциях и интоксикациях в цитоплазме гетерофилов встречаются серо-голубые участки в виде хлопьев или бляшек, так называемые тельца Дёле (Dohle). Это остатки базофильных участков цитоплазмы раннего периода развития клетки.
Количество специальных гранулоцитов резко возрастает в начальной стадии большинства инфекционных болезней («нейтрофильная фаза борьбы»).
В. АГРАНУЛОЦИТЫ
IV. Лимфоциты
Лимфоциты являются типичными агранулоцитами, так как не содержат никакой характерной зернистости в цитоплазме, 8а исключением изредка попадающихся отдельных азурофильных верен. Клетки лимфоцитов округлые, с круглым или овальным ядром, которое окружено или очень узким (малые лимфоциты), или более широким (средние и большие лимфоциты) поясом цитоплазмы. Лимфоциты птиц и амфибий (лягушка) часто встречаются с зафиксированными в момент передвижения псевдоподиями.
Диаметр малых лимфоцитов от 4,5 до 6,5 (л, средних от 6,5 до 10 р. и больших от 10,0 до 18,0 jx.
Цитоплазма лимфоцитов базофильна; при окраске по способу Паппенгейма имеет сетчатое строение, а окрашенная раствором Гимза — гомогенна. Цвет— от бледноголубого у больших и средних лимфоцитов до синего у малых. Вокруг ядра заметна светлая, так называемая перинуклеарная зона. Последний признак помогает диференцировать больших лимфоцитов от не имеющих этой зоны моноцитов. В некоторых (преимущественно малых, иногда средних) лимфоцитах в цитоплазме встречаются в очень небольшом количестве азурофильные зёрнышки (2—8). Крайне редко эти зёрна бывают очень крупными (до 2 ji в диаметре).
Цитоплазма малых лимфоцитов иногда видна лишь с одной стороны ядра в виде очень узкого, едва заметного ободка (форма "серпа"). В некоторых клетках и этот серп незаметен, и тогда малый лимфоцит имеет вид «голого ядра».
Вообще по отношению к цитоплазме ядро лимфоцитов велико, форма его круглая или овальная, особенно правильная у малых лимфоцитов. Часто встречаются ядра с односторонним вдавлением, придающим ядру форму боба (ридеровская форма ядра). Крупные лимфоциты иногда имеют ядро менее правильной формы — угловатое, с выступами или вдав-лениями. В патологических случаях встречаются лимфоциты с неправильной лопастной формой ядра или расчленение ядра может напоминать сегментированные ядра специальных гранулоцитов.
В строении ядра лимфоцитов характерно наличие темноокрашивающихся, неясноочерченных больших глыбок базихроматина, со слабыми просветами между ними. Иногда это чередование тёмных глыбок с тонкими просветами придаёт ядру некоторое сходство с рисунком колеса, спицами которого служат светлоокрашивающиеся участки (А. Н. Крюков и др.). У малых лимфоцитов тёмные глыбки базихроматина настолько сливаются, что структуру ядра установить трудно.
Ядро больших лимфоцитов более рыхлое и менее интенсивно окрашивающееся. В ядре крупных лимфоцитов имеются не всегда ясно заметные 1—2 ядрышка.
Лимфоциты содержат липазу и, повидимому, принимают известное участие в кишечном пищеварении (Синельников). Их базофильная, содержащая некоторое количество нуклеотидов цитоплазма, продуцирует значительное количество иммунных тел (Догерти и Вайт) (Dougherty, White) (1945 г.).
Наконец, лимфоциты участвуют в образовании, из белков плазмы крови, трефонов (Хрущев).
Лимфоциты составляют большинство клеток белой крови у крупного рогатого скота (50—60% всех лейкоцитов), свиней (45—60%), овец (55—65%), коз (40—50%), кур (45—65%) и кроликов (50—65%). У этих животных имеется так называемый лимфоцитарный профиль крови. У собаки и лошади количество лимфоцитов в крови меньше; там превалируют специальные гранулоциты. Однако и у этих животных число лимфоцитов остаётся довольно значительным (20—40% от всех белых кровяных телец).
Количество лимфоцитов в крови молодых животных больше, чем в крови взрослых (за исключением первых дней после рождения). У низших позвоночных количество лимфоцитов может быть относительно очень велико.
В клинике лимфоцитоз встречается в конце благоприятно протекающего инфекционного заболевания («лимфоцитарная фаза выздоровления»). Лимфоцитоз характерен для лимфатической лейкемии, встречается при инфекционной анемии у лошадей и некоторых других заболеваниях.
V. Моноциты
Моноциты — большие клетки крови (от 10,0 до 20,0 µ в диаметре), большей частью округлой, иногда неправильной формы, с хорошо выраженной цитоплазмой, имеющей мельчайшую азурофильную зернистость, и большим, часто эксцентрически расположенным ядром с бухтообразными вдавлениями и лопастями.
Мелкая азурофильная вернистость цитоплазмы почти не видна у моноцитов сельскохозяйственных птиц.
Цитоплазма моноцитов слегка базофилъна, голубовато-серого или пепельно-серого цвета («цвета сигарного дыма») при окраске раствором Гимза п свинцово-серого пли грязноспнего цвета при окраске по способу Паппенгейма. Перинуклеарной зоны нет или она выражена очень слабо. По Крюкову, особенности окраски цитоплазмы моноцитов зависят от того, что преобладающая в ней пара-плазма методом Паппенгейма красится частью в синий цвет, частью в розовый, причём в некоторых клетках превалирует синяя субстанция при почти полном отсутствии розовой, в других обилие розовой субстанции оставляет явственный, своеобразный . отпечаток на морфологическом облике клетки, придавая её протоплазме фиолетово-синий или серо-фиолетовый тон.
У птиц цитоплазма моноцитов серовато-голубая и мало отличается от цвета цитоплазмы лимфоцитов.
Азурофильная зернистость моноцитов хорошо выявляется при окраске по Паппенгейму и с трудом, только при длительной и очень хорошей окраске, — по Гимза. Зернистость розово-красная, очень мелкая, пылевидная.
Ядро сравнительно велико, обычно образует выступы (лопасти) и бухтообразные углубления. Оно имеет очень нежную, тонкую структуру. Ядро моноцитов амблиохроматично (бледно окрашивается), с широконитчатой, мягкой, «облачносливающейся», неравномерной хроматиновой сетью. Интенсивность окраски ядра моноцитов гораздо слабее, чем у лимфоцитов.
Моноциты — это типичные макрофаги И. И. Мечникова. Они захватывают и переваривают остатки распавшихся клеток, попадающие в кровь, инородные частички, в том числе некоторые бактерии, и играют значительную роль в образовании иммунных тел.
В моноцитах имеется протеолитический фермент типа катепсина.
Нормальное количество моноцитов в крови млекопитающих и птиц колеблется в пределах от 2 до 8%. Моноцитоз (повышенное содержание моноцитов) наблюдается в первую фазу выздоровления при большинстве случаев инфекционных болезней («моноцитарная защитная фаза, или фаза преодоления»), при инфекционной анемии лошадей, протозойных заболеваниях и большинстве других инфекционных болезней. По Н. М. Николаеву, однако, моноцитоз при заболеваниях далеко не всегда благоприятный признак, знаменующий собой начало выздоровления.
Плазматические клетки (клетки раздражения)
Плазматические клетки (клетки раздражения) характеризуются одним общим для них признаком — резкой базофилией цитоплазмы (ультрамариновый цвет). Иногда в цитоплазме видны вакуоли. Эта весьма немногочисленная группа клеток имеет полифилетическое, главным образом лимфоцитоидное или миэлоидное происхождение. В соответствии с этим, ядро клеток Тюрка имеет структуру, соответствующую структуре ядер тех клеток, из которых они возникли, но окраска его всегда относительно темнее. Форма ядра — круглая или овальная. Правильные глыбки хроматина придают ядру пятнистый и несколько пикнотический характер.
Вокруг ядра обычно хорошо заметна перинуклеарная зона, периферический же слой цитоплазмы окрашен в интенсивносиний (ультрамаринового оттенка) цвет. Форма клеток — овальная, иногда сильно вытянутая или полигональная, реже круглая. Расположение ядра обычно эксцентричное. Структура цитоплазмы волокнистая или комковатая.
Из плазматических клеток могут возникнуть резко отличные от них по виду дегенеративные формы. В протоплазме плазматических клеток, утрачивающей базофильность, появляются крупные эозинофильные гранулы, вначале имеющие игольчатую форму. Ядро пикнотизируется, и клетка распадается. Гранулы распавшихся клеток, проникшие в соединительную ткань, называются русселевскими тельцами.
Плазматические клетки в крови млекопитающих в заметных количествах встречаются только при патологии. Ими характеризуется так называемая «пёстрая картина крови». У сельскохозяйственных птиц они имеются и в нормальной крови (у кур 0,1 %, по Лебедеву, у гусей до 1,5%, по Домрачеву).
КЛАССИФИКАЦИЯ БЕЛЫХ КЛЕТОК КРОВИ ПО Н. М. НИКОЛАЕВУ
Описанная до сих нор классификация лейкоцитов основывается, главным образом, на их морфологии, так как современное состояние наших знаний о физиологии белых кровяных клеток недостаточно для составления обоснованной функциональной классификации лейкоцитов. Однако некоторые попытки в этом отношении имеются.
Одной ив таких попыток является классификация лейкоцитов Н. М. Николаева. Он рассматривает зернистость белых кровяных клеток как момент, тесно связанный с их функцией. Хорошо выраженная зернистость — признак высокой реактивности клеток мезенхимы.
Соответственно этому основному положению, Н. М. Николаев выделяет пять групп лейкоцитов:
![]()
I группа А
эндотелий
I группа моноцит
гистиоцит
миэлобласт
![]()
I группа Б
промиелоцит
миелоцит
I группа юный нейтрофил (гетерофил)
I группа В
Палочкоядерный нейтрофил (гетерофил)
II группа { сегментоядерный нейтрофил (гетерофил)
III группа { эозинофил одно- и двуядерный
IV группа { базофил
![]() V группа А
V группа А
Лимфобласт
Клетки раздражения
V группа Эритрогоний
Эритробласт
Нормобласт
V группа Б
Лимфоцит
К первой группе Н. М. Николаев относит исходные (материнские) клетки и близкие к ним; ко второй — сегментоядерные и нейтрофилы (зрелые микрофаги); к третьей — клетки фагоцитировавшие (Н. М. Николаев считает, что зёрна эозинофилов — это остатки поглощённых эритроцитов); к четвёртой — дегенеративные клетки и, наконец, к пятой — так называемые синтетические клетки, образующие или гемоглобин (эритробласты), или глобулин (лимфоциты).
В качестве принципиально новой клеточной формы среди агранулоцитов Н. М. Николаев выделяет так называемый гистиоцит или микромоноцит. При окраске раствором Гимза он может быть отличен от лимфоцитов по таким признакам (цитируем по Н. М. Николаеву):
|
Лимфоцит Узкий пояс протоплазмы Интенсивная базофилия протоплазмы Гомогенная или грубозернистая протоплазма Азурчфильные зёрна редки или единичны Темнофиолетовая окраска ядра Густое пикнотрчное ядро, иногда глыСчатое Перинуклеариая зона вокруг ядра |
Гистиоцит Более широкая протоплазма Светлоголубая или сероватая протоплазма Сетчатая или вакуолизиро-ванная протоплазма Большей частью азурофиль-ные зёрна Более светлая окраска ядра Более равномерное, менее плотное ядро Перинуклеарная зона часто отсутствует |
В ряде случаев такое выделение гистиоцитов начинает проникать в практическую медицину и ветеринарию.
РАЗМЕРЫ ЛЕЙКОЦИТОВ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Видовые различия в величине белых кровяных телец незначительны. Ниже приводится таблица диаметров эритроцитов, лейкоцитов и кровяных пластинок у сельскохозяйственных и лабораторных животных.
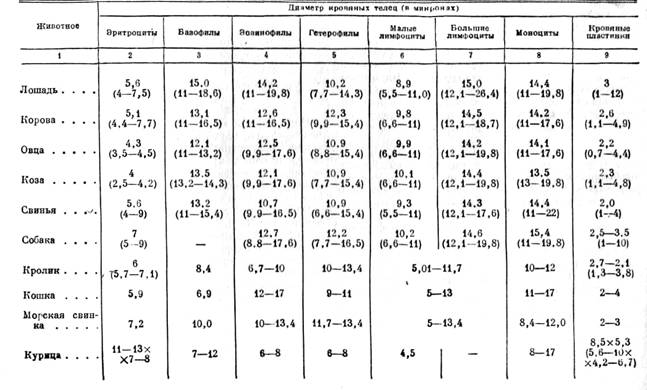
Размеры (диаметры в µ) кровяных клеток у лабораторных и сельскохозяйственных животных
ЛЕЙКОЦИТАРНАЯ ФОРМУЛА И ЛЕЙКОЦИТАРНЫЙ ПРОФИЛЬ КРОВИ
Количественный и качественный состав крови зависит от функционального и патологического состояния организма. Сложные биохимические и физиологические изменения, происходящие в организме при различных патологических состояниях, изменяют функциональное состояние гемопоэтической системы, а стало быть, и состав крови. Эти влияния изучены в совершенно недостаточной степени. Решающая роль здесь, несомненно, принадлежит изменению характера обмена веществ в самих периферических тканях. Некоторое представление об этом даёт схема взаимоотношений кроветворных органов с железами внутренней секреции, в основном заимствованная у Н. М. Шустрова и X. X. Владоса (1930 г.) (рис. 3).
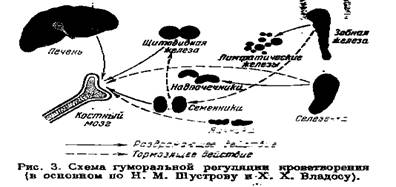
Известно также влияние нуклеиновокислого натра, вызывающего лейкоцитоз. Несомненное, хотя и мало изученное, влияние на кроветворение оказывает активная реакция ретикулоэндотелиальной системы тканей внутренней среды.
При патологическом раздражении вегетативной нервной системы наблюдаются две фазы лейкоцитоза: 1-я фаза — лейкоцитоз с миелоидной тенденцией, сочетающийся при болезни с усилением лихорадочного состояния, обмена веществ, ускорением распада белков, повышением содержания сахара и падением содержания холестерина в крови и ацидозом,—преимущественно симпатикотония; 2-я фаза — лейкопения с лимфатической тенденцией, эозинофилия, ослабление лихорадки и обмена веществ, замедление распада белков и вообще явления, противоположные наблюдающимся в 1-й фазе, преимущественно ваготония. Влияние блуждающего нерва на эозинофилню установлено с достаточной достоверностью, так же как влияние симпатического на нейтрофилию.
На гемопоэз влияют следующие основные гуморальные факторы:
а) продукты
распада красных кровяных телец и,
возможно, лейкоцитов — стимулирующе;
б) щитовидная железа — стимулирующе;
в) печень — стимулирующе;
г) половые гормоны: андроген — стимулирующе— и эстроген — угнетающе (это установлено только для кролика);
д) ряд витаминов группы В, прежде всего фолеивая кислота, — стимулирующе;
е) токсины микроорганизмов, особенно патогенных, и продукты их распада (действие неоднозначное и диференцированное по отношению к различным видам кровяных клеток);
ж) гуморы: ацетилхолин, адреналин (влияние мало изучено);
з) антианемический фактор желудка — стимулирующе (П. А. Троицкий и др.);
и) гормон селезёнки — угнетающе.
Под воздействием этих гуморальных факторов количественный и качественный состав крови своеобразно меняется. Однако следует иметь в виду, что картина крови oтражает (и то не всегда прямо) функциональное состояние лишь кроветворных органов, а не организма в целом. При этом токсины, действующие, например, на нервную ткань, могут не оказывать существенного влияния на систему кроветворения, и наоборот. Очевидно, при различных заболеваниях состав крови может быть одинаковым, и, наоборот, одно и то же заболевание, в зависимости от функционального состояния кроветворных органов, может дать различные картины крови. Поэтому Е. Фрейфелъд (1948 г.) считает, что при пользовании лейкоцитарной формулой нужно руководствоваться следующими положениями: 1. Так как кроветворная система является для организма очень важным органом, то необходимо знать, как она функционирует, точно так же, как необходимо знать функции сердца, почек и т. п.
2. По крови мы устанавливаем функциональную диагностику кроветворной системы, некоторые функции которой вам известны, других же мы не знаем.
3. Ввиду того, что некоторые заболевания дают резко выраженный различный морфологический состав крови возможно исследованием крови исключить одно заболевание и высказаться в пользу другого.
4. Так как кровяные клетки постоянно сменяются новыми, то в случаях, когда они выявляют морфологически действие токсина (токсичность лейкоцитов, различные степени созревания — сдвиги), можно легко проследить, когда действие токсина прекращается, и, наоборот, выявить его, как только оно появляется.
5. При заболеваниях, дающих определённую кривую лейкоцитарной формулы, отклонения от неё указывают на неправильность течения или осложнения.
6. Реакция кроветворных органов не есть выражение иммунобиологического состояния всего организма, поэтому при диагностике нужно считаться с тремя возможностями:
а) реакция кроветворных органов соответствует реакции всех остальных органов, — тогда картина крови аналогична общему состоянию больного;
б) реакция кроветворных органов вследствие их недостаточности слабее реакции остальных органов, — тогда по картине крови можно поставить значительно худший прогноз, чем он окажется в действительности; в) несмотря на тяжесть заболевания, функция кроветворной системы превышает реактивную способность остальных органов, и тогда картина крови будет лучше, чем состояние «больного».
К изменениям картины белой крови относятся:
а) увеличение и уменьшение общего количества лейкоцитов;
б) изменение процентного соотношения отдельных видов лейкоцитов;
в) изменение морфологических свойств (дегенеративные и регенеративные формы) отдельных клеточных
элементов в крови
Лейкоцитарная формула учитывает качественные сдвиги состава белой крови, лейкоцитарный
профиль Мошковского и качественные и количественные сдвиги
Подсчёт лейкоцитарной формулы производится по окрашенным мазкам крови. При подсчёте методом меандра, подвижным столиком постепенно сдвигают мазок крови по отношению к объективу микроскопа под прямыми углами зубчатой линии (напоминающей по ломаной древнегреческий орнамент — меандр).
Схема этого движения следующая (рис. 4).

Однако такой метод подсчёта не учитывает того, что клетки крови распределяются в мазке неравномерно: на периферии мазка относительно больше гра-нулоцитов, а в глубине — лимфоцитов. Поэтому, особенно у животных с лимфатическим профилем крови (корова, свинья, кролик, коза, овца, курица), лучше «прорезать» полем зрения микроскопа всю толщу мазка, как это видно на следующей схеме (рис. 5).
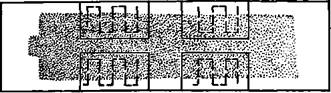
Рис.5 Подсчет лейкоцитов при сплошном прохождении полем зрения мазка
Подсчитав от 100 до 200 (для научных целей и более) лейкоцитов, попавших в поле зрения микроскопа, получаем гемограмму.
Для гемограммы сельскохозяйственных животных целесообразно заменить название «нейтрофилы» на «специальные гранулоциты», или «гетерофилы», так как у некоторых животных вместо нейтрофильной имеется псевдоэозинофильная зернистость. Для крупного рогатого скота и овец возможно разделение лимфоцитов на малые и большие. Профессор Н. М. Николаев предлагает выделить гистиоцитов (микромоноцитов). Значительное количество исследований крови показывает, что лейкоцитарная формула крови у некоторых видов животных (например, кролика) имеет значительные индивидуальные вариации. Поэтому А. А. Заварзин предложил различать животных с филогенетически устоявшейся формулой крови и с филогенетически лабильной формулой. В раннем онтогенезе лейкоцитарная формула более лабильна, с выраженным лимфоцитарным профилем.
Возрастные изменения картины крови у сельскохозяйственных и лабораторных животных
Рациональное применение метода изучения картины крови, как для клинических, так и для зоотехнических и общебиологических целей, требует обязательного знания онтогенеза сосудистой крови, гематологических исследованиях нельзя не учитывать особенностей возрастной эволюции организма, качественного своеобразия каждой стадии онтогенеза: характерных для данной стадии развития черт тончайшей структуры клеток и тканей, соотношения тканей в целом организме, показателей напряжения и динамики процессов, характеризующих отдельные функциональные системы, а также свойственных данному возрасту соотношения и характера гуморальной и нервной регуляции жизненных процессов.
Так как всё это отражается не только на картине нормальной крови, но и на тех изменениях, которые вызываются в ней различными патогенными факторами, то знание онтогенеза крови имеет большое значение и для правильного чтения гемограмм.
Кроме того, ряд авторов указывает на наличие сезонной изменчивости состава крови (Кудряшев и др.).
С другой стороны, онтогенез белой крови, образование лейкоцитов в красном костном мозгу и других местах гемопоэза и содержание в сосудах организма различных форм лейкоцитов не может не отражать последовательной смены нейрогуморальных регуляций и изменений общего функционального состояния организма в онтогенезе. Поэтому индивидуальная эволюцпя белой и красной крови весьма интересна, как своеобразный, легко определяемый, хотя, повидимому, не всегда адэкватный, показатель («сигнализатор>) функциональной и регуляторной индивидуальной эволюции всего организма в целом.
ОНТОГЕНЕЗ БЕЛОЙ КРОВИ У ОСНОВНЫХ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Крупный рогатый скот
Возрастные изменения белой крови у крупного рогатого скота были первоначально изучены И. С. Токарем (1938 г.) для красной степной породы, а затем автором настоящего атласа (В. Н. Никитин. 1946 г.) для остфризской породы. Данные И.С. Токаря представлены в таблице 9.
Свои основные выводы И. С. Токарь формулирует следующим образом:
1. Число лейкоцитов с возрастом уменьшается; максимальное количество (11228 в 1 мм 3) наблюдается в возрасте 3—9 месяцев.
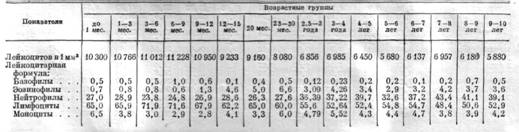
2. Картина белой крови существенно изменяется с возрастом. В молодом возрасте наблюдается лимфоцитоз, причём количество лимфоцитов доходит до 71.9%.
3. Отмечается большая связь процента лимфоцитов с ростом и развитием животных, а также и с половым созреванием.
4. Число нейтрофилов с возрастом увеличивается. Особый скачок нейтрофилов наблюдается с момента отёла.
5. Эозинофилы с момента полового созревания значительно повышаются, с л актированием наблюдается понижение.
6. Для базофилов и моноцитов не обнаружено определённой закономерности в изменении с возрастом.
7. Юные и миелоциты обнаружены только у молодых животных».
И. С. Токарь не дифференцировал в итоговой таблице различные стадии зрелости нейтрофплов и слил в одну группу анализы крови телят моложе 1 месяца.
8. Н. Никитин (1946 г.) исследовал все возрастные периоды крупного рогатого скота остфризской породы, дифференцируя степени зрелости ядра нейтрофилов.
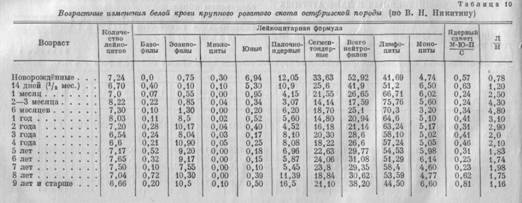
Основные закономерности онтогенеза белой крови у крупного рогатого скота сводятся к следующему:
1. Количество лейкоцитов в онтогенезе изменяется сравнительно мало. Можно отметить слабо выраженный максимум между 3 и 12 месяцами и некоторое уменьшение количества лейкоцитов у взрослых животных.
2. Возрастные изменения в содержании базофилов и моноцитов незначительны и не имеют определённой закономерности.
3. Возрастные изменения количества эозинофилов выражены весьма чётко. В раннем периоде онтогенеза, до 1 года, наблюдается гипоэозинофилия. В годичном возрасте — резкое, скачкообразное увеличение, сохраняющееся на том же высоком уровне в течение всего дальнейшего онтогенеза.
4. Общей закономерностью для онтогенеза нейтрофилов (гетерофилов) является стремительное падение их содержания в самом раннем возрасте (от 52—53% у новорождённых до 17—18% у трёхмесячных телят) и медленный, но непрерывный подъём их количества в дальнейшем (до 38—39% у девятилетних и более старых животных).
5. Миелоциты встречаются только в самом раннем онтогенезе, юные довольно типичны для раннего возраста (6,94% у новорождённых и 5,3% у двухнедельных телят).
Количество палочкоядерных форм сравнительно велико у очень молодых телят (12,05% у новорождённых и 10,9% у двухнедельных телят). От месячного до двухгодичного возраста оно находится в минимуме (от 3,07 до 4,52%), затем несколько нарастает и, наконец, резко увеличивается в позднем онтогенезе (11,39% для восьмилетних коров и 16.5% для коров 9 лет и старше).
6. Возрастные изменения количества лимфоцитов представляют собой обратное отражение изменений нейтрофилов.
7. Наблюдающуюся у коров зозинофилию далеко не всегда можно считать патологической.
В возрастных изменениях белой крови лошади наблюдаются, по данным В. С. Любановой (лаборатория В. Н. Никитина, 1947 г.), те же основные закономерности, что и у крупного рогатого скота. Разница только в абсолютных числах гетерофилов и лимфоцитов, присущих лошадям, и в других переломных периодах онтогенеза. Это видно из диаграммы (рис. 10) и таблицы 11.
К числу особенностей онтогенеза белой крови лошади следует причислить:
а) ааметное снижение количества лейкоцитов к старости;
б) слабый лейкоцитоз новорождённых;
в) наименьшее количество нейтрофплов в возрасте 6 месяцев, т. е. несколько позднее, чем у коров;
г) сравнительно короткий период гипоэозинофилии в раннем онтогенезе.
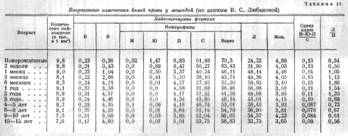
Свиньи
Онтогенез белой крови у свиней исследован В. Н. Никитиным и М. К. Камышанской. У свиней также наблюдаются общие для всех млекопитающих
закономерности возрастных изменений крови. Главная особенность онтогенеза белой крови у свиней состоит в том, что падение и последующий подъём нейтрофилов приходятся на очень ранний период онтогенеза. Повидимому, это связано со скороспелостью организма свиней, усиленной зоотехническим подбором.
Онтогенез белой крови у свиней крупной белой породы виден на диаграмме (рис. И) и таблице 12.
Кроме того:
а) В отличие от лошади и крупного рогатого скота, у свиней в начале онтогенеза количество лейкоцитов понижено (5,04 тыс. в 1 мм3 крови у новорождённых). Оно достигает нормы, характерной для взрослых, к 2-месячному возрасту.
б) Количество базофилов на всех стадиях онтогенеза у свиней заметно больше, чем у других сельскохозяйственных животных (кроме птиц).
в) Гппоэозинофилия очень раннего онтогенеза у свиней длится до 2-месячного возраста и выражена относительно слабо.
г) Минимальный уровень нейтрофилов наблюдается у поросят трёхнедельного возраста. На этот же период падает максимальный уровень лимфоцитов.


Кролики
По данным М. К. Камышанской (лаборатория В. Н . Никитина), онтогенез белой крови кроликов близо к к онтогенезу лейкоцитов у других сельскохозяйственных животных. Главную его особенность состав ляет резкое преобладание лимфоцитов почти на всех этапах индивидуального развития кроликов (кроме новорождённых) (табл. 12а).
ОБЩИЕ ЗАКОНОМЕРНОСТИ ОНТОГЕНЕЗА БЕЛОЙ КРОВИ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Сопоставляя кривые возрастных изменений белой крови крупного рогатого скота, лошадей, свиней и кроликов, можно установить в онтогенезе белой крови всех этих животных некоторые общие закономерности.
Одной из таких основных общих закономерностей является изменение количества нейтрофилов: высокое содержание нейтрофилов в крови новорождённых сменяется быстрым падением его в первые дни жизни и нарастанием числа нейтрофилов в более позднем возрасте.
Обратную картину представляют изменения лимфоцитов.
У лошадей, с их нейтрофильным профилем крови у взрослых особей, такие изменения ведут к двойному перекресту кривых нейтрофилов и лимфоцитов: одному почти в самом начале онтогенеза, другому — в более позднем возрасте.
В картине крови крупного рогатого скота, с его лимфоцитарным профилем крови в стадии стабильного роста, второй перекрест не наблюдается, хотя у старых коров количество нейтрофилов становится очень близким к количеству лимфоцитов. То же самое наблюдается у свиней и кроликов. Общность динамики возрастных изменений отношения лимфоциты (Л) / нейтрофилы (Н) независимо от ее абсолютной
величины, хорошо видна на следующей диаграмме (рис. 12).
По вопросу об основных причинах возрастных изменений нейтрофилов и лимфоцитов пока можно высказать только некоторые предположения. Так, первоначальное богатство крови новорождённых ней-трофилами и быстрое падение их количества в первые дни жизни можно объяснить, например:
а) Явлениями синкаингенеза (Кон — Франк). По этой теории, гормоны, образующиеся в материнском организме, поступая в кровь при плацентарном кровообращении, вызывают у плода такие же изменения, как и в организме матери; к явлениям этого порядка надо отнести, например, увеличение надпочечников, увеличение матки, набухание грудных желез и некоторые другие явления, отмечаемые у новорождённых (А. Ф. Тур).
К этим же синкаингенетическим явлениям можно отнести и резкий нейтрофилёз новорождённых. В дальнейшем, с удалением из организма новорождённого гормонов матери, картина крови всё более определяется собственными гормонами и их соотношением у молодого организма. Отсюда резкое падение нейтрофилов в первые дни после рождения.
б) Нейтрофилёз новорождённых можно рассматривать как приспособление к защите организма от заражения в условиях, когда в первые дни в крови новорождённого имеется лишь очень мало антител (В. Н. Никитин). Дальнейшее падение нейтрофилов и обогащение крови лимфоцитами можно связать:
а) с повторением филогенеза белых кровяных телец на начальных стадиях онтогенеза. Лимфоциты, как наименее диференцированная и древняя форма белых кровяных телец, превалируют на ранних этапах онтогенеза;
б) с значительным развитием и активностью тимуса в раннем онтогенезе;
в) с высокими потенциями роста в равней онтогенезе; лимфоциты, по многим данным, играют известную роль в усвоении и синтезе белков.
За подъёмом количества лимфоцитов происходит
их медленное падение у всех четырёх видов животных. Другой общей закономерностью является нахождение в крови в раннем онтогенезе незрелых форм нейтрофилов. Это ведёт к сдвигу ядра нейтрофилов влево. Он хорошо выражен у крупного рогатого скота и менее заметен у лошадей и свиней.
Вероятно, вначале онтогенеза кроветворные органы функционируют ещё не в такой степени, чтобы нейтрофилы (так же как и другие формы лейкоцитов) успевали в них созреть. Функциональная полноценность миэлопоэтической системы достигается только значительно позднее.
С той же позиции можно рассматривать и некоторый сдвиг влево ядра нейтрофилов у старых животных. Здесь это — проявление уже недостаточной полноценности кроветворной системы. Более того, это своего рода физиологическая раздражённость миэлопоэза, вызванная уже нарушающимися нейро-гуморальными регуляциями в организме.
Третью общую (резко выраженную у всех исследованных животных) закономерность представляет гипоэозинофилия в ранней молодости.
ОСНОВНЫЕ БИОЛОГИЧЕСКИЕ ЗАКОНОМЕРНОСТИ ИЗМЕНЕНИИ БЕЛОЙ КРОВИ
При физиологически нормальном состоянии организма, кроветворные органы выбрасывают в сосудистую кровь достаточно зрелые формы лейкоцитов. В этом случае гемопоэз происходит под влиянием, в общем, довольно стабильных и хорошо сбалансированных нейрогуморальных факторов, обеспечивающих оптимальные условия для полноценного функционирования костного мозга, лимфатических узлов и т. д. Стабильность кроветворения и соответствующая ей стабильность уровня распада белых кровяных телец в организме определяют постоянство состава сосудистой белой крови и общего количества в ней лейкоцитов. Однако постоянство это весьма относительно: в составе белой крови сельскохозяйственных и, особенно, лабораторных животных даже в норме наблюдаются довольно заметные вариации («филогенетически менее устоявшаяся кровь», по А. Заварзину). Особенно вариабильна кровь таких лабораторных животных, как крысы, мыши, морские свинки. Менее изменчива кровь кролика.
Возможно, что на размах колебаний лейкоцитарной формулы сельскохозяйственных и лабораторных животных влияет также и то, что физиологическая норма у них встречается, в сущности, сравнительно редко. Помимо малоучитываемых патологических состояний, почти все сельскохозяйственные и лабораторные животные в той или иной степени инвазированы (гельминтозы, протозоозы и арахно-энтомозы). По мнению академика К. П. Скрябина, «...нигде в мире нельзя встретить ни одной головы крупного рогатого скота, ни одной овцы и лошади, свободной от паразитических червей» (1937 г.).
Наконец, необходимо иметь в виду наличие у сельскохозяйственных животных порою довольно значительных породных различий состава белой крови (а в известной степени и красной). Так, у шаговых пород лошадей лейкоцитарная формула имеет лимфо-цитарный профиль, а у скаковых нейтрофильный. Кровь рысистой лошади занимает промежуточное положение. Считается, что лимфопитарный профиль с наличием эозинофилии характеризует собой ваготоническое состояние, в противоположность выраженному нейтрофильному, свойственному симпатикотонии. Сухая конституция восточной лошади характеризуется симпатикотоническим комплексом, и ему чаще отвечает нейтрофильный профиль, в то время как тяжеловозы с рыхлой конституцией находят выражение ваготоничности в лимфоцитарном профиле.
При патологических процессах изменяется характер нейрогуморальных влияний на кроветворение. На гемопоэтическую систему воздействуют токсины, продукты распада и метаболизма бактерий, специфические антитела и продукты изменившегося катаболизма.
Отсюда двойное влияние на кроветворение в условиях патологии. С одной стороны, раздражённый костный мозг выбрасывает в кровяное русло не только зрелые, но и не закончившие свой цикл развития («вызревания») формы лейкоцитов (сдвиг лейкоцитарной формулы влево, появление в крови лимфобла-стов, монобластов, увеличение числа плазматических клеток).
С другой стороны, особенно при сильных интоксикациях, действие яда непосредственно сказывается на образующейся клетке: появляются дегенеративные формы белой и красной крови.
Основными биологическими закономерностями в изменениях белой крови при патологии являются следующие:
I. Слабые раздражения вызывают лишь функциональные изменения лейкоцитарной картины; средние раздражения, действуя через лейкопоэтические органы, являются формативными; сильные — влияют также на образование отдельной клетки; самые сильные раздражения действуют угнетающе, ослабляя центральные и разрушая периферические клетки кроветворных органов.
II. В большинстве инфекционных процессов первыми реагируют на раздражение нейтрофилы, затем моноциты и в последнюю очередь лимфоциты; различие инфекционных картин крови зависит от взаимного сдвига (во времени) этих трёх фаз и от колебания степени реакции отдельных групп, а также от появления наряду с ними более редких форменных элементов.
По отношению к общему количеству лейкоцитов слабое, умеренное и, в значительном большинстве случаев, сильное раздражение (интоксикация) вызывает нарастание лейкоцитов (лейкоцитоз); при очень сильных, «запредельных» раздражениях, кроветворение подавляется и возникает лейкопения.
Гораздо важнее, конечно, учитывать дифференцированные изменения отдельных групп лейкоцитов. Лейкоцитарная формула при подавляющем большинстве инфекционных заболеваний претерпевает три последовательные фазы изменений.
В начале б о л е з н и наблюдается нарастающий лейкоцитоз, нейтрофилия с резким регенеративным сдвигом ядра влево (до юных форм и даже до миэлоцитов), анэозннофилия, лимфо- и монопения. Это — нейтрофильная фаза борьбы.
Во второй стадии развития болезни, при кризисе и начинающемся выздоровлении, лейкоцитоз постепенно уменьшается, снижается количество нейтрофилов и ослабляется сдвиг ядра влево, появляются эозинофилы (восстановление эозинофилии), несколько увеличивается количество лимфоцитов и резко нарастает число моноцитов (моноцитоз). Это — моноцитарная защитная фаза, или фаза преодоления.
В третьей стадии болезни, при продолжающемся и заканчивающемся выздоровлении, наблюдаются: дальнейшее постепенное падение общего числа лейкоцитов, эозинофилия, резкий лимфоцитоз, несколько уменьшающийся моноцитоз, уменьшение числа нейтрофнлов и постепенное возвращение сдвига ядра до нормы. Это — л и м ф о ц и т а р-н а я фаза, или фаза выздоровления.
Наблюдающиеся при типичном течении болезни фазы изменения лейкоцитарной формулы можно видеть из смены трёх последовательных гемограмм лошади (заимствовано у А. В. Синева):

На следующей диаграмме (рис. 13) показан типичный ход изменений лейкоцитарной формулы крови в течение всей болезни.
Повышенное количество эозтгаофилов иногда удерживается долго после выздоровления. Так, ио данным А. Ф. Дорофеева (1928—1929 гг.), перипневмония у крупного рогатого скота оставляет по себе длительный след на лейкоцитарной формуле, и резко выраженная эозинофилия сохраняется до 3—6 месяцев после клинически установленного выздоровления.
При ряде заболеваний личная смена картин белой крови может существенно изменяться. Так, при крупозной пневмонии у лошадей и коров вначале имеется период лейкопении с уменьшенным количеством нейтрофилов. Затем—резко выраженный нейтрофильный лейкоцитоз. Хорошо выраженная лейкопения наблюдается при инфлуэнце лошадей, паратифе телят и чуме свиней. Интересно, что в то время как чума свиней связана с резко выраженной лейкопенией (всего 2—3 тыс. лейкоцитов в 1 мм3 крови), близкая к ней по начальным клиническим признакам рожа сопровождается не менее сильно выраженным лейкоцитозом (до 25 тыс. лейкоцитов в 1 мм3 крови). Таким образом, диференциальный диагноз рожи и чумы свиней значительно облегчается подсчётом количества лейкоцитов в сосудистой крови.
При гельминтозах, особенно у лошадей, собак и крупного рогатого скота, и при роже свиней наблюдается резкая эозинофилия (до 45—50 % эозинофилов). Гораздо менее выраженная, но всё же отчётливая эозинофилия наблюдается и при некоторых кожных заболеваниях, например, при различных формах экземы, при акариазе у собак.
Наличие значительных, характерных для отдельных болезней, специфических отклонений от типичной биологической реакции кроветворных органов всегда нужно иметь в виду при клиническом диагнозе. Кроме того, эта биологическая реакция даже и в обычных случаях далеко не стандартна.
РАННИЕ ФОРМЫ ОНТОГЕНЕЗА ЛЕПКОЦИТОВ
Для нормальной сосудистой крови характерно наличие в ней более или менее зрелых форменных элементов. В органах же кроветворения, например, в красном костном мозгу, наряду с различными клетками, находящимися на более поздних стадиях вызревания (миэлоцитами, юными, палочкоядерными и сегментоядерными формами), можно найти и самые ранние, исходные для пролиферации и развития, клеточные формы лейкоцитов. Сюда принадлежат (в порядке последовательного вызревания): а) гемоцитобласты (лимфоидоциты, миэлобласт— последний не во всех теориях лейкогенеза является синонимом гемоцитобласта); б) лейкобласты и в) промиэлоциты. В лимфопоэтической системе лейкобластам соответствуют г) лимфобласты.
Кроме того, в красном костном мозгу локализуются гигантские полиморфноядерные клетки д) мегакариоциты, из которых, по видимому, возникают кровяные пластинки млекопитающих. В очень редких случаях (при некоторых заболеваниях, например, хроническом миэлозе) мегакариоциты попадают в сосудистую кровь.
а) Гемоцитобласты (лимфоидоциты) — большие (12—20 µ диаметром) круглые или овальные клетки с базофильной тонкой цитоплазмой и крупным округлённым нерасчленённым, компактным ядром исключительно нежной структуры.
Окружность ядра иногда с бухтообразными углублениями. Хорошо заметна очень равномерная, сетчато-зернистая, без сколько-нибудь заметных сгущений (глыбок) сетка базихроматина. По А. Н. Крюкову, «ядро построено из тончайших примитивных нитей хроматина, отличающихся необыкновенной равномерностью калибра и окраски и дающих чрезвычайной правильности сплетение. Точки перекрещивания нитей ядерной сети воспринимаются как вернистость, откуда создаётся впечатление о нежнейшем микрогранулярном строении ядра. Вследствие малого содержания нуклеина в хроматиновых нитях, ядро получает лептохроматическую окраску, без ясного разделения субстанции ядра на хроматин и оксихроматин. В ядре имеются две-три (до шести) нуклеоли, красящиеся в синий цвет, переходящий в слегка красно-фиолетовый. Нуклеоли имеют характер округлых и более продолговатых образований; некоторые из них являются лишь псевдонуклеолями и представляют собой промежутки в хроматиновой сети ядра, через которые просвечивает синяя субстанция цитоплазмы».
Структура ядра гемоцитобластов может быть охарактеризована и иначе, как имеющая нежную сетчато-зернистую структуру, бледно окрашивающуюся благодаря малому количеству базихроматина (малой насыщенности ядра диффузно распределёнными нуклеотидами). Поэтому в ядре нет чередования отдельных участков с конденсацией бази- или оксихроматина (хроматина и парахроматина).
Нежную структуру поверхности ядра гемоцитобласта сравнивают то с нежной рябью на поверхности воды от слабого дуновения ветра (Крюков), то с поверхностью шагреневой кожи (Паппенгейм), то, наконец, с нежным нитчатым облачком.
Здесь необходимо подчеркнуть, что описание ядерных структур является правильным только по отношению к той искусственно полученной при фпкса-которая отражает коагулировавшие коллоиды ядра. Как показала исследования П. В. Макарова (1946—1948 гг.), покоящееся ядро при жизни не обладает никакими предсуществующими структурами, кроме ядрышка. Нити в ядре и цитоплазме, постулируемые А. Н. Крюковым, следует рассматривать, как артефакты, возникающие при фиксации.
Цитоплазма имеет различную степень базофилии; чем больше развита цитоплазма, тем она бледнее, т. е. менее базофильна. Периферия её окрашивается темнее, вокруг же ядра образуется более слабо окрашивающаяся перинуклеарная зона, иногда приобретающая розоватый оттенок. У тонкопротоплазменных клеток эта зона отсутствует.
Изредка в цитоплазме встречаются азурофильные гранулы.
Гемоцитобласт полипотентен и может развиваться, в зависимости от нейрогуморальных факторов, в направлении миэлопоэза, лимфопоэза, монопозза и еритропоэза.
По Аринкину, в пунктате костного мозга содержится от 1,0 до 1,4% мизлобластов (т. е. гемоцитобластов и последующей формы диференциации в направлении миэлопоэза — лейкобластов).
б) Лейкобласты — промежуточная клеточная форма между полипотентными гемоцитобластами и достаточно чётко диференцированными в сторону грануло-цитопоэза промиэлоцитами и миэлоцитами. Ряд исследователей сближает лейкобласты с миэлоцитами и не выделяет их в самостоятельную промежуточную форму (Максимов, Крюков, Заварзин, Нагели).
По Паппенгейму, лейкобласт характеризуется:
а) ядром, имеющим структуру, приближающуюся к миэлоциту, и иногда содержащим ядрышки;
б) базофильной, несколько более широкой, чем у гемоцитобласта, цитоплазмой, иногда содержащей тёмные, азурофильные зёрнышки, порою очень обильные.
Н. Д. Стражеско и Д. Н. Яновский следующие образом характеризуют особенности лейкобластов:
«Мы под лейкобластом понимаем лимфоидную клетку, ядро которой теряет специальную лимфоидоцитарную структуру. Это не значит, что она должна быть выраженно миэлоцитарной. Эта лимфоидная клетка имеет ядро более грубой структуры, чем ядро лимфоидоцита. Правильное густое сплетение тонких нитей заменяется более грубым сплетением тяжей хроматина с небольшими просветами парахроматина. В ядре части из этих клеток ещё сохраняются ядрышки. Протоплазма не резко базофильна. Ядро значительно отличается от ядра миэлоцита, где плотные участки хроматина чередуются с более светлыми участками, образуя пёструю мизлоцитарную структуру. Так как приобретение определённых биологических свойств идёт вместе с морфологическим созреванием клетки, то потеря специальной лимфоидоцитарной структуры ядра, огрубение его, должны служить доказательством дифрренциации не только морфологической, но и биологической, т. е. приобретения или потери клеткой ряда свойств, как, например, исчезновение полипотентной возможности, т. е. способность развиваться в сторону только лимфоидных или миелоидных клеток, появление ферментов и т. п. В таком понимании лейкобласт будет действительно соответствовать лимфобласту лимфатического ряда. Как раз лейкобласты наиболее трудно отличить от лимфобласта по морфологическим признакам».
Н. Д. Стражеско и Д. Н. Яновский считают, что лейкобласты, уже начавши диференцироваться в направлении гранулопитов ещё сохраняют возможность превратиться, при известных условиях, в моноциты, так же как и лимфобласты. Следующая ступень их развития — промиэлоциты — уже не обладает этой потентностью и может диференцироваться только в гранулоциты.
А. Н. Крюков, не считая возможным выделить лейкобласт в самостоятельную переходную клеточную форму, объединяет, как синонимы, гемоцитобласты, лимфоидоциты и миэлобласты. Н. Д. Стражеско и Д. Н. Яновский объединяют гемоцитобласт и его последующую стадию — лейкобласт — названием миэлобласт.
Выделение лейкобласта в самостоятельную клеточную форму характерно для многих умеренных унитаристов, по отрицается как сторонниками крайнего монизма, так и сторонниками полифилетизма.
в) Промиэлоциты — дальнейшая форма диферен-циации гемоцитобласта и лейкобласта в гранулоциты Она характеризуется:
1) выраженной миэлоцитарной структурой ядра с чередованием ясно выраженных тёмных и светлых участков;
2) отсутствием ядрышек в ядре;
3) появлением азурофильной, уже достаточно обильной, зернистости, которая впоследствии превращается, в зависимости от условий развития, то в эозинофильную, то в гетерофильную, то, наконец, в базофильную зернистость;
4) дальнейшим увеличением массы цитоплазмы Ядро проявляет уже некоторую тенденцию к образованию выступов и вдавлений.
Зёрна дают более или менее выраженную оксидазоположительную реакцию.
Промиэлоциты составляют 0,8—1.4% клеток в пунктате костного мозга (Аринкин).
г) Лимфобласты (макролимфоциты) — такие же промежуточные ступени в направлении развития гемоцитобласта в лимфоциты, как лейкобласты по отношению к миэлоцитам. Лимфобласты трудно отличимы от лейкобластов.
Это большие клетки, с несколько более широким, чем у гемоцитобласта, поясом базофильной цитоплазмы. Их диаметр колеблется между 10 и 15 µ, иногда и более. В отличие от лейкобласта, имеющего несколько дымчатую цитоплазму, у лимфобласта она ясно-голубая.
Большое, имеющее небольшие выступы и впадины, ядро окрашивается бледнее, чем у средних лимфоцитов. Его структура гораздо нежнее, чем у последних (более рыхлая), хотя, в отличие от гемоцитобластов, в нём становятся ясно заметными светлые и тёмные поля. Ядро содержит ядрышки (от одного до трёх), не так резко отграниченные, как в ядре лимфоидоцита.
В цитоплазме ясно выражена светлая перинуклеарная зона, и это служит добавочным отличием лимфобласта от лейкобласта. Базофилия цитоплазмы выражена гораздо слабее, чем у гемоцитобласта. Цвет цитоплазмы — ясноголубой.
Другим, весьма важным, отличием лимфобласта от лейкобластов является их разная оксидазная реакция: у лейкобластов она положительна, у лимфобластов отрицательна.
Лимфобласты находятся в периферической крови только при некоторых патологических состояниях, особенно при лимфатической лейкемии.
д) Гигантские клетки красного костного мозга — мегакариоциты.Мегакариоциты (и их родоначальная, не всеми признаваемая форма — мегакариобласты) представляют гигантские клетки костного мозга, имеющие размер в 20—40 µ и более.
Ядро мегакариоцитов обычно резко полиморфное, образует сложные изгибы (конволюты) и лишь очень редко имеет более простую форму, слегка напоминающую форму ядра моноцитов. Иногда встречаются мегакариоциты с кольцеобразными, сложно извитыми ядрами.
Ядро мегакариоцитов, по Крюкову, «в общем бедно хроматином, дающим явственную сеть из тонких и более толстых нитей с утолщениями в виде частиц, лежащих на разном расстоянии друг от друга. В ядре могут находиться многочисленные нуклеоли».
В ядре мегакариоблаетов видны нуклеоли.
Цитоплазма гигантских клеток костного мозга весьма объёмиста, окрашивается базофильно, мутна. У более зрелых форм в цитоплазме находятся многочисленные мелкие азурофильные зёрнышки.
Особенностью цитоплазмы мегакариоцитов является наличие в ней очень большого количества цент-риоль (по А. Максимову, нескольких сотен).
От цитоплазмы зрелых мегакариоцитов постоянно отделяются небольшие комочки, содержащие азуро-фильную зернистость. Это явление, а также наблюдающийся параллелизм между изменением количества мегакариоцитов в костном мозгу и изменением количества кровяных пластинок в сосудистой крови дали Райту (Wright) основание рассматривать кровяные пластинки как продукт распада мегакариоцитов. В костном мозгу содержится около 0,2% мегакариоцитов (Аринкин).
Мегакариоциты никогда не встречаются в периферической крови физиологически нормального организма. Довольно часто их находят при лейкемиях (преимущественно малые, бедные цитоплазмой, клетки). Н. Д. Стражеско и Д. Н. Яновский считают их не мегакариоцитами, а мегакариобластами.
ГЕНЕЗИС КРОВЯНЫХ КЛЕТОК
ТЕОРИИ КРОВЕТВОРЕНИЯ И ФИЛОГЕНЕТИЧЕСКИЕ И ОНТОГЕНЕТИЧЕСКИЕ ОТНОШЕНИЯ РАЗЛИЧНЫХ ФОРМ ЛЕЙКОЦИТОВ
Картина крови теснейшим образом зависит от функционального состояния органов кроветворения, отражая в той или иной степени происходящие в гемопоэтической системе изменения. Поэтому для правильного истолкования картины крови большое значение имеет знание закономерностей кроветворения в норме и в патологических условиях и, прежде всего, генеза кровяных клеток.
К сожалению, до сих пор ещё нет единой, безупречно обоснованной теории кроветворения. Известно, что у беспозвоночных и низших позвоночных животных, не имеющих кроветворных органов и дифференцированных белых кровяных телец, все элементы крови возникают из индиферентных мезенхимных клеток. Нечто подобное наблюдается и на самых ранних стадиях эмбриогенеза млекопитающих. Однако и в филогенезе (у более высоко организованных животных) и в онтогенезе (на более поздних стадиях эмбриогенеза) отношения значительно усложняются. Возникают специальные органы кроветворения (миэлоидная и лимфоидная системы), диференцируются белые кровяные тельца. Генезис клеток крови у высших животных оказывается гораздо более сложным, чем у животных примитивных. Он изучен далеко не в достаточной степени. В настоящее время существуют три основные теории генезиса клеток крови:
а) монистическая, или унитаристическая, монофилетическая теория (Усов, Вайденрайх, Максимов, Немилов, Доминичи, Феррата, Хлопин, Мясоедов, Алёшин и др.);
б) дуалистическая теория (Эрлих, Негели и др.);
в) триалистическая теория (Ашоф и др.). Особую теорию представляет собой умеренный
унитаризм пли монофилетический дуализм (Крюков, Паппенгейм, Стражеско и Яновский, Тимофеевский), объединяющий унитаризм и дуализм, устраняя их крайние положения.
Некоторые основные положения признаются почти всеми исследователями. Считается, например, установленным, что в эмбриональном кроветворении недиференцированные клетки мезенхимы в процессе диференциации (и пролиферации) образуют первые островки кроветворения, продуцирующие все основные виды клеток крови и в дальнейшем развивающиеся в кроветворные органы. При этом возникают две кроветворные системы: миэлоидная (костный мозг) и лимфоидная (лимфатические узлы и селезёнка). Обе системы диференцируются уже в эмбриогенезе и образуют при физиологически нормальных условиях различные клетки крови: миэлоидная—кровяные пластинки, красные кровяные тельца и гранулоциты, а лимфатическая — лимфоциты. Диференциация мезенхимальных элементов в этих тканях зашла так далеко, что превращение одной ткани в другую (или обратное развитие) в обычных физиологических условиях встречается редко (унитаристы) или исключено (дуалисты и триалисты). Морфологическая диференциация сочеталась здесь с диференциацией биохимической; одно из самых ярких проявлений последней — наличие оксидаз в миэлоидных элементах и отсутствие этих ферментов в лимфоидных клетках (только моноциты иногда слабо оксидазо-положительны). Различны и условия внутриклеточного пищеварения у гранулоцитов и моноцитов; у первых пищеварительные ферменты действуют в слабощелочной среде, а у вторых в слабокислой. Окислительный обмен у гранулоцитов сдвинут в сторону гликолиза, а у агранулоцитов гликолиз выражен очень слабо.
Однако у взрослых организмов, как в кроветворных органах, так и вне их, сохраняются и мало изменившиеся индиферентные мезенхимные клетки. Под влиянием особых физиологических и патологических воздействий они могут развиваться в любые кровяные клетки — как миэлоидного, так и лимфоидного ряда.
При физиологически нормальных условиях лейкопоэз обеспечивается не этими индиферентными мезенхимными клетками и даже не первыми формами их начинающейся диференциации — миэлобластами и лимфобластами, а размножением ещё более диференцированных форм — миэлоцитов и соответствующих им по степени зрелости форм агранулоцитов.
Основная проблема теории кроветворения состоит в том, до какой стадии созревания сохраняется у взрослого позвоночного полипотентность клеток кроветворной ткани, иначе говоря, насколько далеко зашла диференциация миэлоидной и лимфоидной тканей.
По представлению унитаристов (Максимов), грань между миэлоидной и лимфоидной тканью очень относительна, резкого обособления между ними нет, и в обычных физиологических условиях возможно образование лимфоидных элементов из мало диференци-рованных элементов миэлоидной ткани и наоборот. Унитаристы считают, что все агранулоциты крови и кроветворных органов (малые и большие лимфоциты и моноциты) являются недиференцировашшми или лишь очень мало диференцированными клетками, которые при некоторых условиях могут развиться в гранулоциты, эритроциты и мегакариоциты.
По А. Максимову, «соотношения между всеми... формами незернистых лейкоцитов в циркулирующей крови гораздо более тесные и простые, они не только происходят из одной общей родоначальной клетки, что, конечно, несомненно, но они и в самой крови и в других тканях могут в известных пределах прямо превращаться друг в друга. По крайней мере, в крови животных... очень часто вообще не удаётся провести резкой границы между ними, а, наоборот, удаётся подобрать непрерывный ряд постепенных переходных форм от малого лимфоцита к большому (где последний циркулирует в крови) и к моноциту, который является как бы наиболее зрелой формой лейкоцитов. Большой же лимфоцит или моноцит, размножаясь делением, могут опять дать начало малым лимфоцитам».
«На основании этой нерезкой отграниченности друг от друга разных
видов незернистых лейкоцитов, они и могут быть объединены... под общим
названием «лимфоциты» или «агранулоциты>. Это всегда в основе одна и та же
равнозначная клетка. Внешний вид её, величина, отношение между массой
протоплазмы и ядра, степень базофильности протоплазмы, отношение к прижизненной
окраске и т. п. могут значительно колебаться, и клетка может соответственно
этому более или менее приближаться то к одному, то к другому из описанных трёх
типов, но внутренние потенциальные её качества при этом остаются неизменными».
«Это учение признаёт, что все вообще лимфоидные элементы в организме по
существу совершенно равнозначны, хотя в гистологическом отношении могут быть
очень разнообразны. Как бы они ни были различны по виду, величине, отношению
между объёмом ядра и протоплазмы, по базофильности последней и т. д., это всё-таки
всегда те же самые индифферентные блуждающие клетки, мезенхимные амёбоциты,
лимфоциты в широком смысле», с очень большой «потенцией развития; малые
лимфоциты, большие лимфоциты, моноциты представляют собою лишь некоторые наиболее
типичные внешние формы, которые может принимать этот элемент в организме... все
эти клеточные типы постепенно переходят друг в друга и связаны промежуточными
формами. То, что описывается под именем гистиоцитов, лейкобластов и т. д.,
также не может считаться определёнными и резко ограниченными клеточными типами.
Сказанное относится затем и к миэлобластам и лимфобластам дуалистов, т. е. к
незернистым родоначальным клеткам миэлоидной и лимфоидной ткани. Они должны
рассматриваться не как две различные клеточные формы, а как один и тот же
индиферентный, мезенхимный амёбоцит, который можно в данных двух случаях по
гистологическому виду назвать просто большим лимфоцитом. Если этот 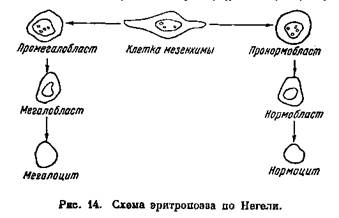 большой
лимфоцит в миэлоидной ткани, развиваясь и диференцируясь, даёт в результате
эритроциты и зернистые лейкоциты, а та же самая клетка в лимфоидной ткани,
размножаясь, производит только себе подобные элементы, то этот на первый
взгляд странный факт объясняется очень просто тем совершенно естественным
предположением, что направление развития лимфоцита в каждом данном случае
зависит всегда от тех внешних условий, в которых он находится. Очевидно, что
внешние условия в миэлоидной и лимфоидной
ткани для одной и той же клетки должны быть совершенно различны и в результате
этого одна и та же клетка и даёт в обоих случаях различные продукты развития»
(Максимов).
большой
лимфоцит в миэлоидной ткани, развиваясь и диференцируясь, даёт в результате
эритроциты и зернистые лейкоциты, а та же самая клетка в лимфоидной ткани,
размножаясь, производит только себе подобные элементы, то этот на первый
взгляд странный факт объясняется очень просто тем совершенно естественным
предположением, что направление развития лимфоцита в каждом данном случае
зависит всегда от тех внешних условий, в которых он находится. Очевидно, что
внешние условия в миэлоидной и лимфоидной
ткани для одной и той же клетки должны быть совершенно различны и в результате
этого одна и та же клетка и даёт в обоих случаях различные продукты развития»
(Максимов).
Унитарная теория кроветворения видна на схеме, заимствованной у Максимова (цветная таблица 64).
Все клетки в этой схеме размещены по линиям, которые показывают их постепенное днференциальное развитие в определённом направлении, начиная от родоначальных индиферентных больших лимфоцитов (1—3) и кончая соответствующими зрелыми формами циркулирующей крови (11, 12, 13, 23, 27, 34). Наиболее сильны позиции унитаристов в области сравнительной гематологии и эмбриологии.
Полифилетические теории опираются, главным образом, на данные клинической диагностики.Дуалисты (Негели, Шридде) резко обособляют миэлоидную и лимфоидную ткани. Они считают диференциацию этих тканей зашедшей так далеко, что возникновение клеток одной системы из родоначальных клеток другой совершенно невозможно ни в физиологических, ни в патологических условиях. Далёким отголоском когда-то существовавшей связи и единства этих двух систем является у взрослых млекопитающих только индиферентная мезенхимная клетка.
Ниже приводится дуалистическая схема кроветворения по Негели (из А. Егорова) (рис. 14, 15, 16).
Широко распространена среди гематологов умеренно унитарная или монофилетически-дуалистическая теория генеза кровяных клеток (Крюков, Стражеско, Паппенгейм). По этой теории, не толькс примитивный гистиогенный элемент (индиферентная клетка мезенхимы), но и следующая стадия его развития — лимфоидоцит (гемоцитобласт) является связующим звеном и общей родоначальной клеткой миэлоидной и лимфоидной систем. Это узкопротоплазменная, лимфоцитоподобная клетка с нежной и тонкой структурой ядра («шагреневая кожа»). Лимфоидоцит (гемоцитобласт), в зависимости от различных
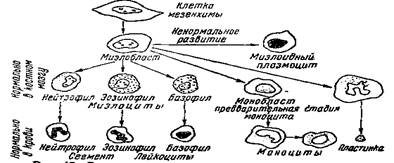
условий развития, может диференцироваться то в лимфобласт (родоначальную клетку для лимфоцитов), то в лейкобласт (родоначальную клетку миэлоидного ряда клеток). Гетерогенное развитие начавшего диференцироваться лимфоидоцита уже невозможно.
Генез крови по теории умеренного унитаризма виден из схем А. Н. Крюкова и Н. Д. Стражеско и Д. Н. Яновского (рис. 17 и цветная таблица 63).
А. Н. Крюков следующим образом характеризует пограничное положение умеренного унитаризма по отношению к унитаризму и дуализму:
«По этой теории, тканевое кроветворение идёт из примитивного
гистиогенного элемента всегда через стадию лимфоидоцита. Лимфоидоцит одинаково
является источником неоплазмы миэлоидной ткани и повсеместных лейкемических
лимфоцитов. Лимфоидоциты выступают на сцену при образовании круглоклеточного
экссудата при воспалении. Они являются также первыми клетками в эмбриональном
кроветворении. Лимфоидоцит обладает способностью диференцироваться в
зернистые лейкоциты, лимфоциты и эритроциты. Эта теория проводит разграничение
меж ду различными лимфондными элементами, различая между ними миэлоидные клетки
и лимфатические в среди тех и других выделяя особые типы —
лимфои-
доцит, лейкобласт или лимфоидоцит с ядром миэло-цита, макролимфоцит, лимфоцит. В то время как унитаризм все различия между этими лимфоидными элементами отрицает и считает их лишь различиями функционального значения, дуализм известным морфологическим различиям придаёт чрезмерное значение, как это существует в вопросе о материнских клетках. Умеренный монофилетизм оценивает эти различия менее высоко там, где они менее выражены, как в вопросе о материнских клетках, и здесь ближе стоит к унитаризму, но приближается к дуализму в вопросе о различиях между мизлобластами и лимфоцитами. Унитаризм считает лимфоцит эмбриональной клеткой, дуализм — высоко диференци-рованным элементом, эквивалентом полиморфному лейкоциту; умеренный унитаризм, признавая малые лимфоциты за зрелые элементы, тем не менее не ставит их на одну доску с полиморфными гранулоци-тами, так как эти последние уже потеряли способность к делению, которая принадлежит лимфоцитам в полной мере. Дуализм противополагает миелоидную ткань лимфоденоидной, унитаризм отрицает обособленность кроветворных систем, умеренный монофилетизм считает что обе ткани суть различные формы развития одного и того же эмбрионального лимфоидоцита, через который они находятся между собою в родственных отношениях». «Лимфоциты по этой теории образуются и в костном мозгу, как особый парамиэлоидный тканевый компонент, первоначально возникший всё-таки из этой же общей материнской клетки, специфицировавшейся в костном мозгу в направлении гранулопластики. При патологических же условиях, как это бывает при сепсисе, развитие лимфоидоцитов в гранулоциты сильно страдает, и тогда созревание может проявляться в сторону лимфопластики, среди скудных миэлоидных элементов обнаруживаются в значительном количестве лимфоциты, что является для костного мозга уже феноменом патологического характера» (Крюков).
Наконец, по триалистической теории кроветворения, имеются три
родоначальные формы кровяных клеток, исходные для возникновения лимфоцитов,
моноцитов и гранулоцитов. Гранулоциты возникают в красном костном мозгу,
лимфоциты — в лимфатической системе и селезёнке, а моноциты в
ретикуло-эндотелиальной системе. Таким образом, по представлению триалистов,
имеются три различные кроветворные ткани — миэлоидная, лимфоидная и ретикуло-эндотелиальная.
Наиболее существенным подтверждением этой теории является обнаружение особой
формы лейкемии — моноцитарной, наряду с ранее известными миэлоидной и
лимфоидной лейкемиями. 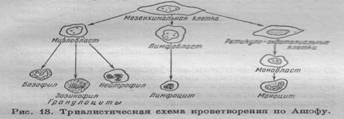
Ниже приводятся схемы кроветворения по триалистической теории (рис. 18 и 19).
Новую концепцию, пытающуюся преодолеть крайности и некоторую догматичность основных теорий кроветворения, выдвигает А. А. Заварзин (1932 г.).
По его мнению, «... такое обилие противоречивых теорий указывает, несомненно, на неправильный подход к самой постановке вопроса. Эта неправильность, по нашему мнению, состоит в том, что авторы множественных теорий пытаются приписать родоначальным формам абсолютную детерминированность, а авторы монистических воззрений — абсолютную лабильность (неустойчивость). Между тем, весьма вероятно, что для камбиальных элементов тканей внутренней среды не существует ни абсолютной детерминированности, ни абсолютной лабильности.
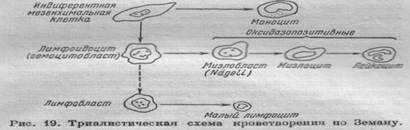
Есть много данных, говорящих о том, что в известных условиях кроветворный камбий детерминирован лабильно. Лабильная детерминированность обусловливает те факты, которые лежат в основе всех множественных теорий кроветворения.
При более резких вмешательствах, при более значительных общих или местных изменениях эта лабильная детерминированность нарушается, и тогда создаются условия для различных перестроек в кроветворении, которые дали повод к обоснованию монистических воззрений».
Последняя концепция представляется нам особенно важной. Она дает возможность преодолеть ограниченность полифилетизма и крайнего монизма. Лучше всего с этой концепцией согласуется современная теория умеренного унитаризма, развиваемая рядом ведущих отечественных гематологов (А.Н.Крюков, Н. Д. Стражеско, Д. П. Яновский, И. А. Кассирский, Г. А. Алексеев, Б. Л. Алёшин, А. Д. Тимофеевский). На позициях этой теории стоит и автор настоящего атласа.
Картина кроветворения, представленная изложенными здесь теориями, наблюдается у млекопитающих. У птиц имеется существенное изменение, сводящееся, во-первых, к тому, что лимфоидная ткань рассеяна по всему организму гораздо более диффузно и настоящие лимфатические узлы встречаются лишь у немногих видов, во-вторых, у них, как у всех позвоночных, имеющих ядерные эритроциты, последние образуются исключительно внутри кровеносных сосудов красного костного мозга. В костном мозгу птиц имеются широкие венозные капилляры (так называемые синусоиды) с чрезвычайно замедленным движением крови. Внутренняя поверхность эндотелия этих сосудов покрыта лежащими в несколько рядов эритробластами, дающими при делении и последующем вызревании зрелые эритроциты. Последние, по мере вызревания, оттесняются образующимися из зритробластов клетками к центру сосуда и затем уносятся током крови. Экстраваскулярно происходит образование красных кровяных телец и у всех остальных низших позвоночных. У рептилий и бесхвостых амфибий эритропоэз локализуется в красном мозгу, у хвостатых амфибий и рыб — в селезёнке.
Зернистые лейкоциты образуются у птиц (и у рептилий и высших амфибий) экстраваскулярно, в самой ткани костного мозга. У низших позвоночных (хвостатых амфибий и рыб) красного костного мозга нет и гемопоэз становится более диффузным, причём нет резкого разделения лимфоидной и миэлоидной тканей. Чрезвычайно мало известно о генезе тромбоцитов у птиц. По 'некоторым данным, они образуются из лимфоцитов, в виде особого самостоятельного вида клеток, по другим — они возникают из эндотелия. У млекопитающих же, повидимому, кровяные пластинки образуются из мегакариоцитов (гигантских клеток) красного костного мозга, путём отшнуровывания цитоплазмы (Райт).
КРОВЯНЫЕ ПЛАСТПНКИ МЛЕКОПИТАЮЩИХ И ТРОМБОЦИТЫ ПТИЦ И НИЗШИХ ПОЗВОНОЧНЫХ
Другим видом форменных элементов крови млекопитающих являются так называемые кровяные пластинки. Это маленькие тельца (от 1 до 3—4 µ в диаметре), с неправильными «рваными» краями и утолщением посредине. Каждая пластинка состоит из гиалоплазмы (гиаломер), образующей основу пластинки, и хромомера — зёрнышек, скопляющихся в её центре или, изредка, разбросанных по гиалоплазме. Гиалоплазма окрашивается по Гимза и по Палпенгейму в голубовато-серый цвет (иногда с ро-вовым оттенком). Зёрнышки хромомера окрашиваются азурофильно, в вишнёвый цвет.
В среднем 1 мм3 крови сельскохозяйственных млекопитающих животных содержит от 200 тыс. до 400 тыс. кровяных пластинок. Однако у отдельных видов имеются значительные колебания количества пластинок в крови. Размеры кровяных пластинок у некоторых животных (кролик, свинья) мало вариируют, у других же (особенно у лошади) вариации в величине весьма значительны: при среднем диаметре в 3 µ, попадаются и очень маленькие пластинки (до 1 µ) и прямо гиганты, достигающие 12 µ.
Кровяные пластинки не являются клетками. По-видимому, это осколки клеток, но происхождение их недостаточно ясно. Вероятнее всего предположение Райта, установившего, что гигантские клетки костного мозга — так называемые мегакариоциты, отшнуровывая выступы своей цитоплазмы, содержащие азурофильные зёрна, образуют кровяные пластинки. Исследования Райта были подтверждены рядом авторов. Однако Максимов считает кровяные пластинки остатками дегенерировавших и вытолкнутых из эритроцитов ядер.
Функциональное значение кровяных пластинок также не может считаться хорошо выясненным. Их способность быстро распадаться, слипаться друг с другом в плотные кучки, с возникновением вокруг таких кучек нитей фибрина при свёртывании крови говорит за активное участие кровяных пластинок в этом процессе. По некоторым данным, кровяные пластинки (а также и эндотелий сосудов) содержат тромбокиназу. Способность кровяных пластинок образовывать, при нарушениях кроветока, плотные сгустки ведёт нередко к закупориванию (тромбу) мелких кровеносных сосудов.
На препаратах кровяные пластинки чаще всего встречаются кучками, причём иногда границы отдельных пластинок исчезают. Опытный исследователь, изучая мазок крови, может отметить тромбопению (очень малое количество кровяных пластинок) и тромбоцитов (повышенное их количество).
Предполагается, что при тромбопении скорость свёртывания крови резко снижена.
В крови птиц и всех низших позвоночных, наряду с эритроцитами и лейкоцитами, встречается третий тип клеток — тромбоциты. Кровяных пластинок у них нет. Каковы филогенетические взаимоотношения между тромбоцитами птиц и низших позвоночных и кровяными пластинками млекопитающих, неизвестно.
Тромбоциты — это овальные (у амфибий веретенообразные) клетки с большими ядрами и сравнительно тонким слоем облегающей их цитоплазмы, которая только на полярных концах клетки скопляется в несколько большем количестве. В цитоплазме нередко видно (на концах клетки) несколько азурофильных зёрен. Иногда цитоплазма вакуолизирована (у гусей очень часто). Цитоплазма очень слабо окрашивается в нежноголубой цвет (иногда остаётся бесцветной). Однако границы её видны очень чётко. Ядро содержит мелкие, интенсивно окрашивающиеся глыбки хроматина.
Размеры тромбоцитов отдельных видов сельскохозяйственных птиц незначительно различаются. Так, у гуся средняя длина тромбоцитов равна 6,8µ, ширина 4,5µ; у курицы — соответственно 8,5 µ и 5,3 µ В 1 мм3 крови имеется от 25 до 70 тыс. тромбоцитов.
Функция тромбоцитов изучена ещё меньше, чем кровяных пластинок млекопитающих. На препарате они нередко собираются кучками, но клеточные границы при этом сохраняются.
По Мевесу, в препарате свежей крови тромбоциты очень быстро подвергаются некробиотическим изменениям. При этом они слипаются кучками, прилипают к стеклу, укорачиваются, но в то же время расширяются (набухают); цитоплазма, скопляясь на одной стороне клетки, образует всё более утончающиеся псевдоподии, превращающиеся в тончайшие лучи. В окружающей плазме вокруг таких тромбоцитов начинают образовываться нити фибрина. Всё это даёт основание предполагать, что функция тромбоцитов близка к функции кровяных пластинок млекопитающих, т. е. они играют существенную роль в свёртывании крови.
Образование тромбоцитов у птиц локализуется в красном костном мозгу. По одним авторам, они образуются из особых родоначальных клеток, по другим — из эндотелия сосудов.
КРАСНЫЕ КРОВЯНЫЕ ТЕЛЬЦА (ЭРИТРОЦПТЫ)
Количественно преобладающей клеточной формой нормальной крови позвоночных животных являются красные кровяные тельца — эритроциты. Обычно количество их в 1 мм3 крови исчисляется миллионами, в то время как кровяные пластинки y птиц и низших позвоночных—тромбоциты) исчисляются в том же объёме крови сотнями тысяч, а лейкоциты — тысячами.
Поэтому на мазках физиологически нормальной крови основной фон составляют густо лежащие друг около друга, окрашенные эозином в яркорозовый или медно-красный цвет эритроциты.
Красные кровяные тельца выполняют в организме исключительно важную функцию — перенос кислорода от лёгких к тканям. Это осуществляется благодаря содержанию в эритроцитах железосодержащего сложного белка — гемоглобина. Обычно в эритроцитах бывает 33% гемоглобина (соответственно 12—17% гемоглобина в цельной крови). Каждый грамм гемоглобина, переходя в оксигемоглобин, связывает 1,34 см3 кислорода, образуя с ним легко диссоциирующее химическое соединение.
Совокупность эритроцитов всей крови животного называется эритрояом. У лошади весом 500 кг эритрон состоит из 436,5 триллиона красных телец, общим объёмом в 14,4 л и содержит 6,76 кг гемоглобина. По мазку крови можно, при известном навыке, составить приближённое представление как о количестве эритроцитов по густоте расположения клеток на равномерно полученном мазке, так и о насыщенности их гемоглобином — по интенсивности окраски (методом Романовского) каждого отдельного эритроцита. Для подсчёта количества красных кровяных телец и для точного определения количества гемоглобина применяют специальные, методы исследования крови. Подробное описание этих методов дано в любом курсе физиологии животных. Картина красной крови при специальной окраске мазка особенно ценна тем, что она даёт возможность распознавать регенеративные и дегенеративные изменения в эритроцитах по разной интенсивности окрашивания их специфическими красками, а также по изменению формы и внутренней структуры эритроцитов.
А. НОРМОЦИТЫ
Картина красной крови физиологически нормального взрослого животного характеризуется безусловным преобладанием зрелых форм красных кровяных телец — нормоцигпов. Сравнительно очень редко среди нормоцитов, окрашенных по методу Романовского в типичный медно-красный цвет, попадаются и незрелые эритроциты — полихроматофилы, окрашенные в переходные цвета от ясно синего, типичного для юной формы, через сине-фиолетовый, до фиолетово-красного цвета, приближающегося к нормальной окраске зрелого эритроцита. Таких форм бывает не более 1—5 на 1 000 зрелых эритроцитов у коров и лошадей и несколько более у свиней, собак, морских свинок и крыс.
Нормоцит млекопитающих (за исключением верблюда и ламы) представляет собой круглую безъядерную, плоскую клетку, с утолщёнными краями и несколько вогнутым центром. Собственно, вернее даже было бы говорить не о клетке, а об остатке клетки, поскольку нормоцит лишён обязательной и важнейшей составной части клетки — ядра. (Поэтому для элементов красной крови млекопитающих лучше применять название «красное кровяное тельце», чем «эритроцит», хотя последнее очень широко распространено и имеет преимущество краткости.)
У верблюда и ламы нормоциты овальны.
В профиль нормоцит имеет вид бисквита. Форму нормоцита лучше представить в виде пластинки или диска с утолщёнными краями. По некоторым новым данным, эритроциты в циркулирующей крови имеют колоколообразную форму («шапочки») с вогнутым центром. На неокрашенном мазке красные кровяные тельца кажутся жёлтыми или зеленовато-жёлтыми, соответственно цвету гемоглобина в очень тонких слоях. Периферическая часть, как содержащая более толстый слой гемоглобина, окрашена интенсивнее.
При окраске по Гимза эритроциты окрашиваются в красивый розово-красный, а при окраске по Паппенгейму — в медно-красный цвет. Так как при этом избирательно окрашивается гемоглобин, то на периферии, в утолщённой части эритроцита, где гемоглобина больше, окраска выражена интенсивнее. В центре окраска несколько менее интенсивна, но, в норме, достаточно заметна. При нарушениях гемоглобино-образования, нормоциты окрашиваются атипично. Иногда резко ослаблена окраска только центральной части красного кровяного тельца. Тогда эритроцит кажется красным кольцом с просветом в центре, — так называемая кольцевая форма. Такие формы особенно типичны даже для физиологически нормальной крови собаки.
В других случаях количество гемоглобина падает настолько сильно, что весь эритроцит (но, конечно, в первую очередь его центр) окрашивается гораздо слабее нормального. Такие эритроциты называются гипохромными, а само явление — гипохромией.
Наконец, возможны случаи, когда эритроциты содержат больше гемоглобина, чем обычно. Такие эритроциты окрашиваются интенсивнее и называются гиперхромными (явление гиперхромии).
При изучении мазка с дополнительным подсчётом количества эритроцитов и определением количества гемоглобина можно установить очень важный показатель насыщенности каждого отдельного эритроцита гемоглобином — так называемый цветной индекс (показатель) крови.
Цветной показатель не может быть определен, даже весьма приближенно, по мазку крови. Казалось бы, интенсивность окраски красных кровяных телец эозином даёт основание для суждения о насыщенности эритроцитов гемоглобином. Однако это далеко не так. Густота окраски эритроцита зависит, кроме фактора интенсивности (концентрации гемоглобина), также и от фактора ёмкости (размеры эритроцита, его толщина). При некоторых анемиях (особенно при микроцитарной гиперхромной анемии) резко изменяется форма красных кровяных телец. Из плоских, относительно растянутых дисков они превращаются в толстые, гораздо меньшего диаметра, тельца. При этом значительно возрастает густота окраски таких, кажущихся более мелкими, эритроцитов. В действительности содержание гемоглобина в таких эритроцитах не изменяется или изменяется в гораздо меньшей степени, чем это представляется при рассматривании их в окрашенных мазках.
.Цветной показатель (У) обозначает не абсолютное содержание гемоглобина в одном эритроците, но некоторую пропорциональную абсолютному содержанию величину. Уровень гемоглобина в кровп дан в условных процентах по Сали. В норме цветной
показатель равен единице (У = 1,0). Число большее единицы указывает на избыток гемоглобина в эритроците (гиперхромия); цветной показатель меньше единицы указывает на пониженное содержание гемоглобина (гипохромия).
Цветной индекс для сельскохозяйственных п лабораторных животных нужно рассчитывать по следующей полной формуле:
NRxHb
J= AHbxR
где: J — цветной индекс; NR — нормальное для данного вида количество эритроцитов в 1 мм3 крови; NHb — нормальный для данного вида животных. Многие авторы считают, что в сосудах животного эритроциты имеют чашеобразную или даже колоколообразную форму (Геле, Вайденрайх, Крюков). Возможные прижизненные изменения

формы эритроцитов представлены на следующей схеме (рис. 20).
По величине эритроциты можно подразделить на собственно нормоциты (для лошади — 5,6 µ диаметром), микроциты и макроциты. Микроциты это эритроциты меньшего, чем в норме, диаметра (для лошади — менее 5 µ), макроциты — большего (7—6 µ). Внутренняя структура эритроцитов почти не выяснена, но самое наличие этой прижизненной структуры кажется весьма вероятным. Иначе было бы непостоянной формы эритроцитов, «теней эритроцитов» понятно наличие их эластичности, нахождение при гемолизе, проникновения в эритроцит трипанозом без выхода из него гемоглобина, несомненно до казанное наличие в нём особых специфически окрашивающихся образований, и т. д. С поверхности красное кровяное тельце отграничено липоидно-белковой мембраной (Крюков, Лепешинская). В какой степени она отдиференцирована гистологически, представляется

ещё спорным. Наличие чётко выраженной оболочки эритроцита защищается Немиловым и Лепешинской.
Под оболочкой предполагается наличие «краевых обручей» — эластических нитей, образующих остов эритроцита (рис. 21).
Весьма вероятно наличие в эритроцитах «внутренних тел», указываемых Максимовым, Арнольдом и др.
Ряд исследователей (в том числе Н. Д. Стражеско) развивают представление об очень сложной прижизненной структуре так называемого «совершенного эритроцита» млекопитающих. Эта, в значительной степени гипотетическая, структура представляется состоящей из:
1. Ядра, остатков ядра или кровяных пластинок (кр. п.).
2. Протоплазмы, состоящей из:
a) радиальной структуры, лишь редко видимой (С);
b) нагромождённой сверху в юном возрасте базофильной субстанции (полихромазия);
c) коркообразной сложной наружной оболочки (М).
3. Архоплазмы, состоящей из:
a) более светлого центрального вещества (ст. т.), соответственно вогнутости («стекловидное тело»);
b) микроцентра (центральное тельце) с соединением (ц); в микроцентре имеются два очень маленьких блестящих зёрнышка;
c) прилежащего, трудно изобразимого, величиною в 1—2 микрона, шаровидного, так называемого «капсулъного тела» (К).
Вряд ли, однако, можно признать такую сложную структуру достаточно экспериментально обоснованной. Более того, имеются высказывания об отсутствии такой сложной структуры в эритроците (Насонов). Не совсем понятно, какие физиологические функции могли бы быть связаны с такой сложной и в значительной степени искусственной структурой красного кровяного тельца (рис. 22).
Эритроциты птиц и низших позвоночных существенно отличаются от красных кровяных телец млекопитающих прежде всего тем, что даже в зрелом состоянии содержат ядра. Кроме того, они гораздо крупнее размером и имеют овальную форму.
Потеря зрелыми формами эритроцитов млекопитающих ядра произошла, вероятно, в процессе приспособления этих клеток к переносу кислорода
Ядерные эритроциты птиц и низших позвоночных являются полноценными клетками с интенсивным обменом веществ и поэтому значительное количество переносимого ими кислорода потребляют сами. Эритроциты же млекопитающих, теряя ядро, резко снижают свой газообмен и, следовательно, мало потребляют переносимый ими кислород. Безъядерные эритроциты, следовательно, более «экономные» переносчики кислорода, чем кариоциты птиц и низших позвоночных.
В мазках крови эритроциты видны иногда тесно наложенными друг на друга («монетные столбики»). Особенно резко эта способность выражена в крови лошади. Очень трудно получить мазок лошадиной крови, где бы эритроциты не образовывали, накладываясь друг на друга, густой сети. Отдельные красные кровяные тельца обычно находятся только на тонком, свободном краю мазка крови лошади.
При медленном подсыхании мазка резко повышается концентрация солей плазмы крови, и в таком гипертоническом растворе эритроциты, отдавая воду, принимают неправильную звёздчатую форму или форму тутовых ягод.
Размеры эритроцитов у различных видов животных значительно вариируют, так же как и их количество. В таблице 14 приведены средние данные о количестве и размерах зрелых эритроцитов у основных сельскохозяйственных и лабораторных животных. Общей закономерностью является обратная пропорциональность между размерами и количеством эритроцитов в 1 мм3 крови.
По В. П. Зайцеву, размер эритроцитов лошади зависит от типа конституции. Так, у астенических лошадей средний диаметр эритроцитов 5,12 µ у мускулярных 5,02 µ и у пикнических 4,9 µ.
В соответствии с этим, и количество эритроцитов, по В. П. Зайцеву, зависит от конституции: в 1 мм3 крови астенических лошадей содержится в среднем

9,97 млн. эритроцитов, у мускулярных 7,51 млн. и у пикников 7,98 млн.
Весьма мало известно о длительности жизни эритроцитов. В отношении безъядерных красных кровяных телец имеются данные о том, что их жизненный цикл составляет 3—4 недели. Они подвергаются фагоцитозу в селезёнке, в расширенных капиллярах её пульпы. Железо их гемоглобина, вместе с частью пиррольных колец гематнна, откладывается в селезёнке в виде железосодержащего пигмента — гемосидерина. Часть гемина, лишившегося железа, попадает в печень и превращается там в жёлчные пигменты. В печени же накопляется обычно и известное количество гемосидерина. Это количество доходит до громадных размеров в патологических условиях, когда происходит усиленный распад эритроцитов и гемоглобина. Образующийся при этом железосодержащий пигмент усиленно накопляется не только в печени и селезёнке, но и костном мозгу и лимфатических сосудах, обусловливая явления их гемосидероза.
Гемосидерин следует рассматривать, как резерв железа и пиррольных колец, который может быть использован для синтеза гемоглобина.
Б. ГЕНЕЗИС ЭРИТРОЦИТОВ
Постоянное новообразование эритроцитов происходит у млекопитающих в красном костном мозгу. Основной, исходной клеткой для развития эритроцитов является лимфоидный эритробласт (по А. Н. Крюкову, прогемобласт, или проэритробласт). Лимфоидный эритробласт является первой ступенью (этапом) диференциации лимфоидоцита (гемоцитобласта) в эритроцит. Из лимфоидного эритробласта возникает непосредственный предшественник эритроцита — эритробласт. За счёт размножения и диференциации эритробласта и происходит, при обычном, физиологически нормальном кроветворении, непрерывное новообразование эритроцитов.
Последовательное образование эритроцита представить в виде следущей схемы:
Схема эритропоэза
Индифферентная мезенхимная клетка.
Лимфоидоцит (гемоцитобласт)
Лимфоидный эритробласт (проэритробласт, прогемобласт)
Полихроматофильный эритробласт
Эритробласт (нормобласт)
Эритроцит (нормоцит)
Стадия лимфоидоцита (гемоцитобласта) может дать, диференцируясь под соответствующими гуморальными влияниями, все виды клеток крови: гранулоциты, агранулоциты и эритроциты. Лимфоидный эритробласт (проэритробласт, прогемобласт) уже начинает диференцироваться в направлении эри-тропоэза и является в этом отношении унипотенциальным.
Лимфоидный эритробласт (проэритробласт). Проэритробласт, эта материнская клетка эритроцитов, по своей структуре ещё весьма близок к родоначальным кровяным клеткам. Это большая клетка (до 20 µ в диаметре у лошади), с крупным, округлым ядром и резкобазофильной цитоплазмой, несколько более широкой, чем у гемоцитобласта. Крупное, почти правильной круглой или овальной формы ядро, при окраске по Романовскому, окрашивается в интенсивный красно-фиолетовый цвет. «Хроматиновая сеть ядра отличается необыкновенной правильностью своего сплетения, равномерностью составляющих сеть примитивных нитей и в то же время нежностью этого сплетения. В большинстве случаев примитивные нити более крупного калибра, чем у лимфоидных материнских клеток, резче красящиеся, и потому ядро получает более тёмную и более насыщенную окраску» (А. Н. Крюков). У более зрелых проэритробластов ядро имеет укрупнённую структуру, что создаёт впечатление зернистости или рубчатости. Иногда внутри ядра встречаются небольшие кругловатые или вытянутые участки, окрашивающиеся в синий или сине-фиолетовый цвет. Это нуклеоли или, вернее, псевдонуклеоли — участки протоплазмы, просвечивающей сквозь структуру ядра.
Цитоплазма проэритробластов окрашивается в интенсивно-синий, с лёгким оттенком ультрамарина, цвет. Она явственно нитчата и делится на две зоны: перинуклеарную, очень узкую, имеющую розоватый оттенок, и гораздо более широкую зону интенсивного фиолетово-синего или ультрамариново-синего цвета при окраске по Романовскому. Переход между этими зонами плавный, но отчётливо заметный.
Эритрпбласт. При дальнейшем созревании лимфоидный эритробласт превращается в эритробласт — клетку, постоянно продуцирующую в костном мозгу млекопитающих эритроциты. В физиологически нормальных условиях лимфоидные эритроциты являются покоящимися, резервными, малодиференцированными клетками, непосредственно не участвующими в текущем эритропоэзе. Только в патологических условиях они получают гуморальный стимул к дифференциации в эритробласты.
Эритробласты в костном мозгу размножаются посредством митотического (кариокинетического) деления. Однако при ряде патологических состояний эритробласты могут делиться и амитотически, но при этом получаются карликовые формы эритроцитов. Итак, начальной клеткой физиологически нормального эритропоэза является эритробласт.
Первоначальная, юная форма эритробласта, так называемый базофильный эритробласт, постепенно переходит в полихроматофильный, а этот — в ортохромный эритробласт.
По мере вызревания лимфоидного эритробласта, структура его ядра становится всё грубее, превращаясь постепенно в типичную для эритробласта радиарную, с большими, тёмными глыбками хроматина, со светлыми между ними промежутками, расположенными по типу спиц в колесе. Цитоплазма становится бледнее, блёклосинего цвета (базофильный эритробласт) с постепенным переходом к сине-фиолетовой окраске (полихроматофильный эритробласт). В дальнейшем, благодаря постепенному накоплению гемоглобина, цитоплазма, окрашенная по Романовскому, имеет сперва жёлто-розовый, а затем типичный для зрелого эритроцита медно-красный цвет (ортохромный эритробласт). На этой стадии созревания ядро эритробласта млекопитающих резко уменьшается и пикнотизируется, радиар-ная структура постепенно исчезает, всё ядро интенсивно окрашивается в вишнёво-фиолетовый цвет и принимает правильную круглую форму. Размеры созревающего эритробласта прогрессивно уменьшаются и, наконец, достигают размеров эритроцита.
Последнее изменение эритробластов (кариоцитов) млекопитающих перед превращением в эритроциты состоит в исчезновении ядра (энуклеации). До сих пор ещё неясно, как это происходит. А. Максимов и ряд других исследователей полагают, что пикнотизированное ядро эритробласта всё более сдвигается к периферии клетки и, наконец, выталкивается из неё. Некоторые думают, что вытолкнутое ядро превращается в кровяную пластинку. Иногда перед выталкиванием ядро, принимая сперва форму розетки и даже сегментов, соединённых между собою мостиками, подвергается распаду на отдельные осколки (кариорексис). При энуклеации эритробласта возникает эритроцит.
Большая часть исследователей считает, однако, что при физиологически нормальных условиях ядро или его осколки растворяются в клетке (кариолизис) (Паппенгейм, Негели, Заварзин).
Наконец, некоторые учёные склонны допустить наличие обоих путей освобождения эритробласта от ядра — и выталкивание его и растворение (Стражеско, Крюков, Вайденрайх, Феррата).
В. МЕГАЛОБЛАСТЫ И МЕГАЛОЦИТЫ
У молодых эмбрионов, а также в патологических условиях постэмбрионального кроветворения проэритробласты диференцируются в эритробласты несколько иного типа, обычно гораздо большего размера, с ядром, сохраняющим в известной степени нежную сетчатость более ранних стадий развития. Такие эритробласты называются мегалобластами, а возникающие из них эритроциты (тоже обычно несколько более крупные и с большим насыщением гемоглобином) — мегалоцитами. Таким образом, в период раннего эмбрионального кроветворения все эритробласты принадлежат к мегалобластам. На более поздних стадиях развития мегалобластнческое кроветворение сменяется нормобластическим и лишь в патологических условиях в костном мозгу снова встречаются мегалобласты, а в крови—мегалоциты.
В физиологической норме всё или почти всё красное кроветворение взрослых животных — нормобластическое, а сосудистая красная кровь представляет собой только нормоцитов. Этот тип эритропоэза сохраняется и в большинстве анемий, и только при пернициозной анемии наблюдается мегалобластический тип кроветворения.
До сих пор являлись неясными филогенетические и онтогенетические взаимоотношения мегалобластического и нормобластического кроветворения. Многие исследователи (Эрлих, Негели, Стражеско и Яновский и др.) резко разграничивают эти два типа клеток, другие же считают, что обе эти формы — лишь крайнее выражение единого типа эритропоэза, и никакой принципиальной разницы между нормобластами и мегалобластами не существует (Крюков, Паппенгейм). По А. Н. Крюкову, «мегалобластическое кроветворение является в результате превалирования клеточного размножения над дифференциацией. Мегалобласты размножаются и созревают, не дифференцируясь, в нормобласты. Кроветворение остаётся на эмбриональной мегалобластической ступени вследствие повышенного регенеративного требования и отравления организма ядом, вызвавшим заболевание. Различие между мегалобластическим и нормо-бластическим кроветворением — только в степени реакции кроветворительной ткани. Нормальный эритропоэз идёт за счёт размножения существующих в костном мозгу эритробластов. Мегалобластический эритропоэз задевает глубже эритропоэтические потенции кроветворительной ткани, и эритроциты возникают из первичных базофпльных лимфоидных клеток, мобилизуемых в случаях крайней надобности и исключительной потребности. Тюрк (Turk) думает, что чем дальше продвигается предсозревание не содержащих гемоглобина эритробластов, прежде чем начнётся образование гемоглобина, тем больший нормобластический характер обеспечен за возникающими эритробластами, так как за это время предсозревания базофильный эритробласт в своём ядре и протоплазме делается меньше, некоторым образом концентрируется. Чем быстрее наступает образование гемоглобина, тем клетка больше и тем больше и нежнее структурировано ядро, т. е. тем ближе клетка к ме-галобласту. Поэтому и первоначальный эритропоэз у эмбриона имеет мегалобластический тип, где примитивные эритробласты являются производными мезенхимных элементов, вторичный же эрптропоэз эмбриона возникает уже из лимфоидных предстадий, имеющих время подготовиться к образованию эри-тробласта. Возможно, что у эмбриона первоначальная усиленная потребность в красных элементах удовлетворяется спешной пролиферацией этих клеток, дп-ференциация же их запаздывает. Только когда развивающийся организм обеспечивается красными клетками в известной степени, становится возможным более надёжный и более стойкий эритропоэз нормобластического типа».
Открытие «фактора вызревания эритроцитов» (образуется в стенке желудка и концентрируется в печени), способствующего переходу мегалобластов в эритробласты, выясняет отношения между последними. Сабин считает мегалобластов нормальной промежуточной формой при эритропоэзе. В его схеме имеются следующие стадии вызревания эритроцита:
Ретикулярная клетка
«Примитивная клетка»
Мегалобласт
Ранний эритробласт
Поздний эритробласт
Нормобласт
Полихроматопит или ретикулоцит
Зрелый эритроцит.
Г. РЕГЕНЕРАТИВНЫЕ И ДЕГЕНЕРАТИВНЫЕ ИЗМЕНЕНИЯ КРАСНЫХ КРОВЯНЫХ ТЕЛЕЦ
Регенеративные изменения в эритроцитах наблюдаются при усиленном эритропоэзе. Полное вызревание эритроцита может при этом несколько нарушаться, давая необычные для нормальной крови формы, и в сосудистую кровь могут поступать не совсем зрелые формы красных кровяных телец, иногда даже самые ранние их предстадии.
Примером регенеративных нарушений нормального типа созревания эритроцитов является преждевременная энуклеация ядра, в то время как цитоплазма, сохраняя присущие юным клеткам нуклеиновые кислоты и не накопив гемоглобина, остаётся ещё базофильной или полихроматофильной. Отсюда появление в сосудистой крови полихроматофильных или даже базофильных эритроцитов. Иногда базофилия цитоплазмы сохраняется в виде небольших участков, пятен, островков на ортохромной поверхности эритроцита. В этом случае появляются базофильно пунктированные эритроциты. Наконец, в созревшем эритроците могут сохраниться остатки ядра — в виде отдельных обломков (азурофильная пунктация, тельца Жоли) или в виде остатков ядерной оболочки (кольца Кабота) (Cabot).
Иногда, наоборот, созревание ядра при стимулированном эритропоэзе отстаёт от созревания цитоплазмы. В этом случае дифференцированная, ортохроматическая цитоплазма окружает незрелое, нежно структурированное ядро. Аналогичное явление получается при ускоренном созревании цитоплазмы.
Все эти, сами по себе патологические отклонения в созревании эритроцитов свидетельствуют об усиленном новообразовании красных кровяных телец, их регенерации, о регенеративных сдвигах в красном костном мозгу.
Однако существуют и такие изменения эритроцитов, которые происходят при угнетении эритропоэза какими-либо вредными воздействиями, ведущими к явлениям дегенерации в красном костном мозгу. Такие дегенеративные изменения выражаются в изменении величины (анизоцитоз), формы (пойкилоцитоз) и окрашиваемости (появление гипер- и гипохромных форм) эритроцитов.
В процессе созревания регенеративные формы, под влиянием вредных воздействий, могут также подвергаться дегенеративным изменениям: в этом случае в эритроцитах возникают смешанные, дегенеративно-регенеративные изменения.
«Относительно истинного значения различных патологических форм красных кровяных элементов не существует полного единодушия во взглядах, и то, что одними признаётся за дегенеративные изменения, другие то же самое считают регенеративными изменениями. Но в общем, исходя лишь из морфологических изменений клетки при прогрессивной дифференциации её, можно составить правильное и последовательное представление и о патологии эритроцитов» (А. Н. Крюков).
Следует иметь в виду, что малые дозы кровяных ядов влияют на кроветворную систему стимулирующе, вызывая, главным образом, явления регенерации (хотя в небольшой степени вызывают и дегенеративные изменения в эритроцитах). При больших дозах кровяных ядов превалируют явления дегенерации, хотя в какой-то степени можно уловить и регенеративные изменения эритроцитов. Только при очень больших дозах кровяных ядов дегенерация совершенно подавляет всякие признаки регенерации.
Полихроматофилы, ретикулоциты и базофильно пунктированные эритроциты. Одним из основных регенеративных признаков красной крови является нахождение в мазке полихроматофилов, ретикулоци-тов и базофильно пунктированных эритроцитов.
При обычной окраске по Романовскому полихромазия выявляется по синевато-фиолетовой или розово-фиолетовой окраске эритроцитов. При суправиталь-ной окраске спиртовым раствором бриллианткрезилголубой зрелые эритроциты окрашиваются в зелёный цвет, а в молодых, недозрелых формах обнаруживаются нежные яркосиние сеточки (ретикулоциты) или отдельные темносиние точки и пятнышки (базофильно пунктированные эритроциты).
Объяснение этому заключается в самой сущности вызревания цитоплазмы. Юная цитоплазма богата нуклеиновыми кислотами, делающими её резкобазофильной (Кедровский). Нуклеиновые кислоты играют большую роль в синтезе белков, в частности, глобулинов в лимфоцитах и, повидимому, гемоглобина в эритробластах. По мере вызревания красных кровяных клеток, базофилия цитоплазмы исчезает, так как количество нуклеиновых кислот в ней уменьшается. Но, при усиленной регенерации, у молодых, поступающих в сосудистую кровь ещё несозревшими, эритроцитов, базофилия частично сохраняется в виде полихромазии, если вся цитоплазма ещё диффузно содержит много нуклеиновых кислот, или в виде ба-зофильной пунктации, если нуклеиновые кислоты сосредоточены в отдельных местах эритроцита.
Количество полихроматофильных эритроцитов в нормальной крови лошади и коровы очень невелико (не более 1—3 на тысячу ортохромных эритроцитов). У всех молодых животных, а у свиней, собак, морских свинок и крыс — также и во взрослом состоянии, полихроматофилов значительно больше. Особенно много полихроматофилов у новорождённых.
При анемиях и некоторых других заболеваниях, при усилении регенерации в красном костном мозгу, количество полихроматофилов в сосудистой крови резко возрастает.
Изменения в эритроцитах, связанные с сохранением остатков распада ядра. а) Красная полихроматофилия, красная пунктация и красная штриховатостъ эритроцитов.
При окраске по Романовскому в эритроцитах можно наблюдать не только базофильную, но и азурофильную полихромазию и пунктацию. В этом последнем случае, благодаря избирательному поглощению азура, эритроцит или целиком окрашен в красно-фиолетовый цвет (красная полихромазпя) или в нём обнаруживается красно-фиолетовая (красная) пунктация в виде отдельных точек или глыбок. Повидимому, азурофильная полихромазия возникает при растворении ядерной субстанции и хроматолизе, а азурофильная пунктация — при распаде ядра на отдельные, очень маленькие глыбки (кариорексис).
Того же происхождения и красная штриховатость эритроцитов, особенно подробно изученная Негели. Она выражается в наличии в эритроцитах отдельных красных (красно-фиолетовых) пятнышек и штрихов.
В норме красная полихромазия и пунктация эритроцитов не встречаются ни у взрослых животных, ни у молодых. Эти явления наблюдаются только при тяжёлых анемических состояниях.
б) Хроматиновая пылинка Вайдеирайха, тельца Говелл—Жоли и кольца Кабота в эритроцитах. Более крупные, сохранившие ещё остаток структуры, ядерные остатки в эритроците дают так называемые тельца Говелл—Жоли, хроматиновую пылинку Вайденрайха и кольца Кабота.
Хроматиновая пылинка Вайденрайха — это последний остаток хроматина ядра в виде тонкого, короткого штриха, зёрнышка, «пылинки», окрашивающейся в вишнёво-красный цвет по Романовскому. Предположение, что это остаток центрозомы, мало вероятно.
Тельца Говелл—Жоли — несколько более крупные ядерные обломки (хроматиновые глыбки), дающие характерные тинкториальные реакции хроматина. По Романовскому они окрашиваются в вишнёво-красный цвет, метилгрюнпиронином — в зелёный. Они окрашиваются также гематоксилином. Наличие этих телец указывает на не вполне закончившийся распад и растворение ядра и, следовательно, на юность красного кровяного тельца.
Кольца Кабота представляют, по-видимому, остатки ядерной оболочки или периферического слоя ядра, образующие замкнутые тонкие кольца, часто в виде цифры 8 или сложных петель. Они окрашиваются азуром в вишнёво-красный цвет.
По А. Н. Крюкову, «они суть патологические продукты, возникающие в результате необычного процесса энуклеации при помощи кариолиза и вакуолизации в противность нормальной центропетальной редукции ядра (Паппенгейм) или выталкиванию ядра (Максимов). Они являются выражением процесса созревания исключительно патологического (Феррата). Но в то же время служат симптомом регенерации и характеризуют недозрелость красного элемента».
Они появляются, и то не всегда, лишь при тяжёлых анемических и лейкемических процессах.
Изменения величины и формы эритроцитов. Изменения величины и формы эритроцитов, особенно грубые и значительные, сигнализируют о дегенеративных изменениях в кроветворной системе.
К таким изменениям принадлежат:
а) Анизоцптоз — появление в сосудистой крови эритроцитов различного, не типичного размера. Формы, большие чем обычный эритроцит (нормоцит), называются макроцитами, формы меньшие — микроцитами. Эритроциты ненормальных размеров возникают иногда из соответствующих материнских клеток — макро- и микроэритробластов. Чаще, однако, они образуются при сморщивании или «набухании» нормоцитов.
Слабый анизоцитоз не обязательно связан с дегенерацией красного костного мозга. Он обычен у очень молодых сельскохозяйственных и, особенно, лабораторных животных.
Сильно выраженный анизоцитоз — всегда признак дегенерации эритропоэтической системы.
б) Пойкилоцитов — появление в сосудистой крови эритроцитов ненормальной, дегенеративной формы. Они могут быть похожи на грушу, гимнастическую гирю, бисквит или имеют своеобразные длинные отростки. Края пойкилоцитов часто неровные, зубчатые или гофрированные.
Выступы эритроцитов могут отшнуровываться, получаются маленькие округлённые обломки — шистоциты. Такое отшнуровыванпе, смотря по своему характеру, называется плазморексисом и плазмо-шизом.
Крупные шистоциты легко смешать с микроцитами. Однако у последних всегда имеется типичная, более бледная окрапшваемость центра клетки, чего нет у шистоцнта.
По А. Н. Крюкову, «пойкилоцитоз можно рассматривать, как дальнейшую дегенеративную ступень анизоцитоза. Пойкилоциты и шистоциты возникают на периферии, но для их происхождения, повиди-мому, требуется продукция костным мозгом весьма мало устойчивых элементов, легко поддающихся изменению под влиянием изменившейся в физико-химическом отношении кровяной плазмы».
в) Анизохромия — ненормально слабая или слишком сильная окрашиваемость эритроцитов (гипо- и гиперцитохромия). В основе их лежат, несомненно, колебания насыщенности эритроцита гемоглобином. В случаях резко выраженной г и п о х р о-м и и центр эритроцита почти или совсем не окрашивается, — возникают так называемые кольцевидные формы эритроцитов. Это особенно характерно для хлороза. Гипохлороз не всегда ведёт к обеднению крови гемоглобином. Иногда (чаще временно) недостаточная насыщенность эритроцитов гемоглобином может компенсироваться повышенным их количеством — поли-цитемией. Гиперхром и я особенно характерна при пернициозной анемии. (Но, например, при ни-кроцитарной гиперхромной анемии гиперхромия связана с утолщением клеток за счёт уменьшения их диаметра.)
г) Полулунные тела. Резко выраженной дегенеративной формой эритроцитов являются полулунные тела. Это бледные, почти бесцветные диски, размером в 10—15 µ а более, ограниченные бледно-фиолетовым слоем. У очень больших полулунных тел этот слой весьма узок. Разорвавшиеся полулунные тела дают серповидные формы, нередко с изогнутыми, как бы извивающимися концами.
По А. Н. Крюкову, эти образования можно рассматривать «как происходящие в результате процесса физиологической инволюции нормальных эритроци-тов, так как они встречаются в значительном количестве у совершенно здоровых индивидуумов». Образование полулунных тел происходит таким образом, «что эритроциты делаются бледнее и метахроматич-ными, далее вакуолизируются, вакуоля в эритроците увеличивается, субстанция же эритроцита окружает эту вакуолю серповидно, причём одновременно сильно увеличиваются размеры инволюционной формы.
В дальнейшем, вследствие разрыва стенки вакуоли, получаются свободные серпы или полулуния. Действительно, подобные формы весьма нередко приходится видеть в крови, не обнаруживающей дегенеративных признаков».
Метгемоглобинемические внутренние тельца в эритроцитах. При дегенерации эритроцитов под влиянием таких кровяных ядов, как фенилгидразин, нитробензол, пиридин, в эритроцитах возникают внутриклеточные метгемоглобиновые образования — так называемые тельца Эрлиха — Гейнца.
При супровитальной окраске метилвиолетом они окрашиваются в интенсивный синий цвет, а нильсульфатголубая окрашивает их в зеленовато-синий цвет. Они избирательно окрашиваются кислыми красками.
Обычно в эритроците находится одно тельце Гейнца. Оно расположено или центрально, или более или менее эксцентрично, а иногда даже выпячивается наподобие клювика за контуры клетки.
Возможно, что тельце образовано не из чистого гемоглобина: кроме метгемоглобина (или, быть может, другого очень стойкого деривата гемоглобина), в нём находили липиды, связанный с белками диаминофосфатид, немного холестерина. С несомненностью доказано присутствие в тельцах железа и пиррольных колец. Тельца Эрлиха — Гейнца химически очень стойки, — они не растворяются в пирогаллоле, воде, эфире и бензине и противостоят действию сапонина.
Общая оценка регенеративных и дегенеративных изменений эритроцитов. Классификация патологических изменении красной крови. А. Н. Крюков делает следующий прекрасный анализ взаимоотношений регенеративных и дегенеративных изменений в красной крови:
«При сколько-нибудь выраженном малокровии картина крови даёт одновременно регенеративные и дегенеративные изменения эритроцитов. Как среди первых, так и среди вторых имеются градации, которые дают возможность судить о степени поражения кроветворительного аппарата, о тяжести заболевания. Наиболее лёгкая степень регенерации определяется полихроматофилией эритроцитов, как выражением ускоренной эритропоэтической деятельности костного мозга. Более повышенная функция сопровождается появлением базофпльной пунктации» (она имеет место и у ядерных эритроцитов птиц. — В. Н.), «далее — эритроцитов с тельцами Жоли, ещё далее — нормобластов. Появление мегалобла-стов будет указанием на гиперфункцию, на энтдиф-ференциацию кроветворительной ткани, когда нормальная регенерация уже становится недостаточной. Точно так же по дегенеративным изменениям эритроцитов можно судить о тяжести процесса. Я вления гипохромии и анизоцитоза будут менее значительными изменениями, чем пойкилоцитоз и шистоцитоз, где деформируются и разрушаются эритроциты. Базируясь на степенях обоего рода изменений в эритроцитах, можно лишь до известного предела строить масштаб анемического состояния, потому что существуют и такие формы малокровии, где, несмотря на незначительные морфологические изменения в крови, тем не менее тяжесть страданий безмерна, благодаря арегенеративному состоянию кроветворительного аппарата. Точно так же стремления конструировать специфические картины эритроцитарных изменений, которые бы характеризовали определённые болезненные формы, оказываются по существу несостоя тельными и возможны только в узких траницах. Так, центральная гипохромия присуща хлорозу, гиперхромия и мегалобластоз — пернициозной анемии. Однако, и другие болезненные процессы могут протекать с этими картинами. Если регенеративная картина крови есть отображение того, что происходит в кроветворительной ткани, то дегенеративные изменения эритроцитов являются в результате воздействия вредных влияний на эритроциты в периферической крови, а также следствием непосредственного отравления кроветворительной ткани».
«...Действительная картина крови есть симптом регенеративного и дегенеративного взаимодействий».
Различные степени патологических изменений крови часто систематизируют в такие четыре категории.
Предварительная стадия. Раздражение (достаточно простого уменьшения количества, благодаря потере крови) вызывает немедленное прямое поступление в кровь резервных клеток (при более сильной степени раздражения — нормальных юных форм) вследствие хемотаксиса или, что более вероятно, вследствие васкулярной гиперемии, быть может регулируемой вегетативной нервной системой, — кратковременная картина крови при раздражении.
1. Скрытая регенерация. Костный мозг в состоянии начинающейся гипертрофии. На периферии обыкновенно находят значительное увеличение числа пластинок и умеренный лейкоцитоз. Выраженные юные формы ещё отсутствуют. В норме непродолжительная промежуточная стадия (2— 3 дня), — псевдоапластические или олигоцитотические картины крови (степень 1а).
2. Простая регенерация. Увеличение полихромазии и её подвидов — признак усиленной, ускоренной деятельности кроветворных органов (гипертрофия костного мозга); при наличии токсического фактора — базофильная пунктацпя, — полихроматическая картина крови (степень IIа).
3. Усиленная регенерация. Резкая полихромазия, юные формы большей величины (макроцитоз), общий анизоцитоз вследствие усиленного образования и поступления в кровь незрелых, отчасти ядросодержащих элементов (нормо- и эритробласты): костный мозг в общем гипертрофирован, в состоянии прогрессирующего красного перерождения (метаплазия); при наличии токсического фактора — обрывки ядер, — нормобластическая картина крови (степень IIIа).
4. Гиперрегенерация. Общее нарушение образования эритроцитов и поступление в кровь действительно новообразованных (изменивших форму или «эмбриональных», богатых гемоглобином, гиперхромных) юных форм: мегалобластов, мегалоцитов и вообще всех дегенеративных форм, указывающее на нарушение нормальной регенерации (прогрессирующая мегалобластическая гиперплазия костного мозга),— мегалобластическая (пернициозная) картина крови (степень IV. а и б).
Н. Д. Стражеско и Д. Н. Яновский утверждают,
что нарушения нормальной регенерации при мегалобластозах нет и что при гиперхромных анемиях типа Бирмера одновременно производятся как нормоциты, так и мегалоциты.
Наблюдающаяся при различных степенях патологических изменений картина красной крови может быть представлена в виде следующей схемы.
НОМЕНКЛАТУРА ГЛАВНЫХ ЭРИТРОЦПТАРНЫХ КАРТИН КРОВИ
Макроцитоз: много больших клеток (макроцитов).
Микроцитоз: много ненормально малых эритроцитов (микроцитов).
Мегалоцитоз: наличие мегалобластов в мегалоцитов.
Анизоцитоз: неодинаковая величина эритроцитов, особенно наличие макроцитов.
Пойкилоцитоз: эритроциты неправильной формы.
Шистоцитоз: наличие в мазке обрывков эритроцитов (шистоцитов).
Полихромазия: присутствие в мазке полихроматофилов вообще.
Базофилия: присутствие в мазке базофильных эритроцитов. То же — для базофильной пунктации,
Олигохромемия, олигохромазия: бледная окраска эритроцитов, определяемая уменьшенным содержанием в них гемоглобина или, гораздо реже, утончением («распластыванием») эритроцитов.
Гиперхромазия: повышенная насыщенность эритроцитов гемоглобином, отсюда — более интенсивная их окрашиваемость. Повышение цветного индекса крови.
Олигоцитемия: уменьшенное количество красных кровяных телец, узнаваемое на равномерном хорошем мазке по их редкому расположению.
Гиперглобулия: повышенное количество клеток. На мазке — очень тесное расположение эритроцитов.

ХАРАКТЕРИСТИКА ОСОБЕННОСТЕЙ КРОВЯНЫХ ТЕЛЕЦ
И КАРТИНЫ КРОВИ СЕЛЬСКОХОЗЯЙСТВЕННЫХ
И ЛАБОРАТОРНЫХ ЖИВОТНЫХ
Лошадь
Эритроциты довольно крупные (средний диаметр 5,6—5,8 µ), мало варьирующие в размерах. У верховых лошадей эритроциты несколько крупнее, у шаговых пород относительно мельче. Количество эритроцитов у скаковых пород заметно больше, чем у шаговых.
Общий вид мазка крови лошади характерен склеиванием эритроцитов в длинные цепочки, которые на толстом мазке пересекаются друг с другом и образуют подобие грубой, неправильной сетки или решётки. Только на конце мазка обычно удаётся найти красные кровяные тельца, лежащие поодиночке.
Полихроматофилы и ретикулоциты встречаются в норме очень редко (менее 1%). Ещё реже наблюдаются пойкилоциты и тельца Жоли.
Наиболее характерной особенностью крови всех однокопытных и, в частности, лошади является структура эозинофилов. Эозинофилы относительно весьма крупны (до 20—22 µ); зёрна их исключительно велики (2—3 µ в диаметре) и до такой степени густо наполняют цитоплазму, что совсем закрывают её. Лишь очень редко бывают видны небольшие участки розовато-серой или серовато-голубой цитоплазмы. Из-под гранул видны отдельные участки ядра; однако из-за массы эозинофильных зёрен форму ядра уловить почти не удаётся. Поэтому невозможно или почти невозможно установить стадию зрелости ядра эозинофила. Лишь изредка у лошадей попадаются сравнительно небольшие эозинофилы с малым количеством крупных зёрен.
Тесное расположение гранул ведёт к их частичному сплющиванию; наряду с круглыми, попадаются зёрна угловатые, иногда сильно вытянутые в длину.
Цвет зёрен в эозинофилах лошади не яркокрасный. а скорее малиново-красный, более бледный, чем у эозинофилов других животных. При хорошей окраске зёрна кажутся удивительно красиво мутно-просвечивающими. Пространства между зёрнами обычно имеют цвет самих зерен, но более бледный. Это просвечивает нижний слой гранул эозинофила.
Базофилы лошади тоже очень крупны (до 18—21µ в диаметре). Они очень легко деформируются. Особенно часто на мазках встречаются базофилы с лопнувшим ядром, окрашивающим всю цитоплазму в вишнёво-розовый цвет. Хорошо сохранившиеся и правильно окрашенные базофилы лошади очень красивы, с сиренево-голубой цитоплазмой и причудливо-извитым, нежной облачной структуры, розово-фиолетового цвета ядром. Гранулы базофилов крупны и вариируют в размерах в одной и той же клетке.
Форма ядра юных, палочкоядерных и даже сегментоядерных специальных гранулоцитов очень часто подковообразна. Сегментоядерные формы чаще всего имеют 3—4 сегмента, реже 2 пли 5.
По Максимову, зернистость специальных гранулоцитов лошади скорее оксифильна, чем нейтрофильна. "Настоящими нейтрофильными специальные зерна..., могут считаться..., например, у обезьяны, собаки, свиньи; у большинства же млекопитающих они должны быть названы амфофильными, так как красятся как кислыми, так и основными красками, причём в одних случаях проявляется большее сродство к первым, а в других — ко вторым. Первый случай встречается особенно часто и касается, например, сравнительно довольно грубых зёрен специальных гранулоцитов кролика и морской свинки, почему они здесь называются также ещё псевдоэозинофильными или более мелких зёрен тех же элементов у быка, овцы, лошади и т. д."
Моноциты имеют в большинстве случаев мало разветвлённое, компактное ядро и относительно бедны цитоплазмой.
Кровяные пластинки, в среднем, довольно велики (около 3 µ) и очень варьируют в размерах. Встречаются и карликовые формы (в 1 µ) и прямо гиганты (достигающие 12 µ). Пределы обычных колебаний размеров кровяных пластинок — от 1,8 до 6,6 µ.
Лимфоциты довольно часто имеют бобовидную форму ядра. Они бедны цитоплазмой. В ней иногда встречаются вакуоли (0,8—5,4%), даже у здоровых животных. Азурофильная грануляция в цитоплазме встречается довольно часто (до 6,1 % у малых и до 14,3% у больших лимфоцитов).
Изредка в норме встречаются плазматические клетки.
Осел
Морфологически кровь осла весьма близка к крови лошади. Диаметр красных кровяных телец колеблется между 5,41 и 6,7 µ, т. е. они несколько мельче, чем у лошади.
Гранулы эозинофилов мельче, чем в эозинофилах лошади. Они не всегда заполняют всю цитоплазму, и между зёрнами часто встречаются голубые цито-плазматические промежутки. Гораздо лучше различима форма ядра.
Базофилы мельче эозинофилов и грануляция их менее крупная, чем базофилов лошади.
Ядро моноцитов компактно, редко образует короткие, округлые лопасти.
Лимфоциты, в противоположность лимфоцитам лошади, имеют чаще всего округлые ядра, без заметных вдавлений или выступов. Иногда, при быстрой фиксации, у лимфоцитов (и у моноцитов) бывают заметны псевдоподии. Цитоплазма скудна (даже и у больших лимфоцитов).
Верблюд
Наиболее характерной особенностью крови верблюда является эллиптическая форма эритроцитов, которые окрашиваются обычно довольно интенсивно, со слабо заметным побледнением в центре. Эритроциты лежат на мазке очень густо, соответственно высокому содержанию их в крови (11 млн. в 1 мм3). Размеры эритроцитов мало варппруют. Их короткая ось равна в среднем 4,0 µ., длинная 7,35 µ. Эллипсоидная (овальная) форма очень правильная. Деформации крайне редки.
Белые кровяные тельца имеют относительно малые размеры, особенно базофплы, диаметр которых в 2 раза меньше, чем у базофилов лошади (всего 8—10,5 µ).
Эозинофилы несколько крупнее, с довольно большими (1,0—1,8 µ.) круглыми гранулами, рассеянными среди ясноголубой цитоплазмы. Форма ядра видна очень хорошо, так же как и у эозинофилов всех остальных приведённых в атласе животных, за исключением однокопытных. Зёрна не «подавляют» ядра.
Ядро моноцитов иногда значительно извито, образует своеобразные петли или довольно длинные лопасти. Изредка в цитоплазме моноцитов попадаются крупные кусочки (тельца), окрашивающиеся как ядро. Возможно, что это — отшнуровавшиеся участки лопастей ядра.
Крупный рогатый скот
Средний размер эритроцитов 5,1 µ (пределы вариаций — от 4,4 до 7,7 µ). Чаще всего размер эритроцитов колеблется в пределах от 4,4 до 5,5 µ.
Среди эритроцитов взрослых животных в норме очень редко попадаются ретнкулоциты. Ещё реже встречаются тельца Жоли. Пойкилоцитоз, полихромазия и эритробласты при физиологической норме или не встречаются совсем, или исключительно редки. Породные колебания количества эритроцитов у крупного рогатого скота относительно невелики (по сравнению, например, с количеством эритроцитов у лошадей).
Эозинофилы коров довольно велики; их цитоплазма наполнена очень ярко красящимися зёрнами, средней или довольно большой величины (0,5 µ). Иногда гранулы лежат в ясноголубой цитоплазме довольно редко. Количество эозинофилов обычно значительно выше, чем v других млекопитающих (в норме 6,0— 8,0% и более).
При депрессии кроветворения в крови коров можно найти карликовые (размером со средний и даже с малый лимфоцит) эозинофилы, цитоплазма которых густо наполнена зёрнами.
Специальные гранулоциты крупного рогатого скота сравнительно велики (больше, чем у всех других сельскохозяйственных млекопитающих). Зернистость слегка оксифильна, выражена хорошо.
Ядро часто значительно расчленено — до 5—8 сегментов.
Бросается в глаза большой размер агранулоцитов.
Моноциты в преобладающей своей части имеют значительно расчленённое, лопастное или лентообразное ядро. Цитоплазма обильна. Мельчайшая азурофильная зернистость около ядра выражена хорошо.
У телят и даже у взрослых коров в крови встречаются большие лимфоциты. Цитоплазма лимфоцитов очень обильна, в ней довольно часто встречаются азурофильные гранулы (5,1 % у малых форм и 12,6% у больших). Очень редко в цитоплазме попадаются весьма крупные азургранулы (до 1,5 µ и более).
Плазматические клетки изредка встречаются в крови и при физиологической норме.
Средний размер кровяных пластинок—около 2,6 µ, Пределы колебаний 1,1—4,9 µ.
В мазке встречается сравнительно много кровяных пластинок, часто слипающихся в большие кучки.
Овца
Очень характерны маленькие, густо расположенные в мазке, эритроциты. Их средний диаметр 4.3µ при обычных колебаниях от 3,5 до 4,5 µ. Однако, встречаются и очень маленькие формы (до 2,5 µ.) и большие (до 7,0 и даже 8,3 µ.). Вообще для овцы обычен небольшой анизоцитоз.
Густота расположения эритроцитов в мазке определяется очень большим их количеством .6—13 млн. в 1 мм3) в крови; это компенсирует их малый размер.
Эозинофилы отличаются большим диаметром (9,9— 17,6 µ); они содержат округлые, иногда неправильной формы, довольно крупные гранулы (0,5—1,0 µ). Зёрна рыхло наполняют обычно хорошо заметную, ясноголубую цитоплазму.
Ядро эозинофилов (как и специальных гранулоцитов) вызревает по кольчатому типу. Поэтому часты формы ядра в виде целого кольца (у палочкоядерных) пли разорванного кольца (у сегментоядерных клеток).
Много эозинофилов со значительной сегментацией ядра (3—5 сегментов).
Базофилы немного меньшего размера, чем эозинофилы (в среднем 12,1 µ при колебаниях в 11,0—13,2µ). Ядро базофилов, особенно при плохой фиксации, очень часто деформировано (лопается). Гранулы крупные, мало варьирующие в размерах, очень темно-окрашивающиеся.
Чрезвычайно характерна большая сегментированность ядер зрелых специальных гранулоцитов. Очень часты формы с 8—10 и более сегментами. Для овцы это норма, а не патологическая форма. Размер специальных гранулоцитов относительно велик, хотя п меньше, чем у крупного рогатого скота. Зернистость по Клинебергеру и Карлу нейтрофильна, по Максимову — с большим сродством к кислым краскам.
Моноциты велики; ядро их сильно расчленено на лопасти, иногда соединяющиеся между собой лишь тонкими мостиками.
У больших и средних лимфоцитов хорошо видна цитоплазма. В ней изредка встречаются вакуоли. Азургранулы в 5,2—6,3% лимфоцитов.
Кровяные пластинки очень малы — 1,9—2,4 µ в диаметре.
Кровь козы по своей морфологии очень близка к крови овцы (особенно лейкоциты). Отличительная особенность эритроцитов козы — их малая осмотическая резистентноетъ. Поэтому мазкп часто получаются со звёздчатой формой красных кровяных телец. Весьма част анизоцитоз. Даже у вполне клинически здоровых животных в крови встречаются пой-килоциты, ретикулоциты, полихроматофилы и изредка даже тельца Жоли. Это указывает на некоторую физиологическую раздражённость эритропоэза, характерную для коз.
Лейкоциты морфологически очень сходны с белыми кровяными тельцами овец.
Свинья
Эритроциты довольно крупные (средний диаметр 5—6 µ). В красной крови свиней в норме встречаются полихроматофилы и ретикулоциты, «кольцевые» формы эритроцитов и даже эритробласты. Сильнее всего это выражено у поросят. Витально гранулированные эритроциты у взрослых евнней встречаются в малом количестве (3—4 на 1 000 нормоцитов); у поросят их количество доходит до 1,1—13,8%.
Белые кровяные тельца не имеют особенно резких видовых признаков.
Базофилы характерны крупными, темноокрашенны-ми гранулами. Эозинофилы невелики, правильной округлой формы. Они имеют яркие, довольно крупные верна (0,5—1,0µ в диаметре). Сегментированность ядра невелика (обычно 2, реже 3 сегмента).
Ядро моноцитов мало расчленено, обычно вытянутой или слегка скрученной формы. Изредка в цитоплазме моноцитов встречаются вакуоли.
Клетки раздражения редки.
В лимфоцитах попадаются азургранулы (4% малых лимфоцитов и 5,8% больших).
Кровяные пластинки относительно малы (около 2,0 µ)-
Количество лейкоцитов в крови довольно велико (10—15 тыс. в 1 мм3).
Собака
Эритроциты, по сравнению с эритроцитами остальных домашних млекопитающих, велики. Их средний диаметр равняется 7µ, при колебаниях от 5 до 9 µ.
На окрашенных препаратах центральная часть эритроцитов и при физиологической норме почти бесцветна («кольцевая» форма).
У взрослых собак в крови встречаются в незначительном количестве (2—3 на 1 000) полихрома-тофильные и витально гранулированные эритроциты и остатки ядра в эритроцитах (тельца Жоли). Очень редко попадаются нормобласты. Имеется слабо выраженный анизоцитоз.
Базофилы крови собак характерны довольно чётко выраженными контурами ядра, правильно круглыми, с резкими границами, сравнительно немногочисленными, неодинаковой величины гранулами и сиренево-розовым цветом цитоплазмы. Базофилов в крови очень мало.
Ядро эозинофплов мало расчленено. Гранулы среднего размера — 0,5—1,0µ в диаметре.
Вызревание ядра специальных гранулоцитов происходит по кольцевому типу.
Ядро моноцитов чаше всего колбасовидное, со своеобразными конволютами на концах. Реже встречаются лопастные формы.
Лимфоциты фиксируются с довольно крупными псевдоподиями. Цитоплазма, даже у больших лимфоцитов, развита умеренно. В ней изредка попадаются вакуоли.
Плазматические клетки встречаются и при физиологической норме.
При некоторых патологических изменениях (инфекциях и, особенно, интоксикациях) в специальных гранулоцитах бывают тельца Деле.
Кошка
Эритроциты довольно велики: средний диаметр 5,9 µ, при крайних колебаниях от 3,2 до 7,5 µ. Довольно часты кольцевые формы красных кровяных телец.
Полихроматофплы и нормобласты в крови при физиологической норме крайне редки.
Базофилы очень велики, с тёмной, розовато-фиолетовой цитоплазмой и крупными, но немногочисленными гранулами. Они очень редко попадаются в крови кошки, и потому Максимов считал, что базофилов у кошки нет.
Эозинофилы средней величины, с округлыми, иногда несколько неправильной формы гранулами. Автор никогда не встречал эозинофилов кошачьей крови с палочковидными гранулами, как это утверждают Вирт, Клинебергер и Карл. Гранулы довольно велики (1,0—1,5 µ). Размеры их значительно варьируют.
Специальные гранулоциты характерны чрезвычайно мелкими зёрнышками в цитоплазме. Контуры ядра на всех стадиях зрелости своеобразно округлены, чётки и изящны. Вызревание ядра происходит, по-видимому, по кольцевому типу.
Ядро моноцитов очень компактно и почти не образует лопастей.
Кровяные пластинки довольно велики (2—4 µ). Пределы колебаний их размеров очень значительны: от карликовых, в 0,8 µ, до гигантских форм, доходящих до 10µ.
Кролик
Эритроциты крупные (в среднем 6,0—6,5—6,8 µ в диаметре). В них очень слабо выражено ослабление окраски в центре. Полихромазия заметно выражена даже при физиологической норме (до 1 %). Ретикулоциты очень часты (до 8% эритроцитов у взрослых животных и 20—80% у новорождённых и в первые месяцы развития). Изредка встречаются нормобласты. Красная кровь вообще весьма лабильна.
Эозинофилы с крупными гранулами (1,5 µ), очень густо расположенными в цитоплазме.
Специальные гранулоциты кролика весьма своеобразны. Их зернистость гораздо крупнее, чем у нейтрофилов других животных, и красится комбинацией красок по Романовскому в яркокрасный цвет, т. е. эозинофильна (оксифильна). Поэтому специальные гранулоциты кроликов получили название псевдоэозинофилов. Форма зёрен неправильно округлая, порою угловатая.
Одинаковая окраска и почти одинаковые размер и форма псевдоэозинофилов и эозинофилов кролика очень затрудняют диференциацию этих клеток. Недавно (1945 г.) Якоби предложен для этой цели следующий специальный метод окраски.
Свежеприготовленный, высушенный на воздухе мазок крови кролика фиксируют спиртом с формалином и затем в течение 5—10 минут окрашивают 0,05-процентным водным раствором
2—6—-дихлорфенолиндофенола (4 части) и 0,5% раствора нейтрального красного (1 часть). К каждым 5 см3 этой смеси прибавляют перед окраской 4 капли перекиси водорода.
После быстрой промывки — высушить фильтровальной бумагой и немедленно смотреть в микроскоп под иммерсией.
При такой окраске протоплазма эозинофилов плотно набита сильно окрашенными большими сферическими гранулами (1,5 и в диаметре); почти каждая гранула имеет тёмную пурпурно-чёрную периферию и немного более светлую внутренность, цвет которой колеблется между темнопурпуровым и грязносероголубым. У псевдоэозинофилов же очень скудные, отдельные темнопурпурные гранулы (диаметром 0,5 µ), скорее овоидной и жезлообразной формы.
Ядра псевдоэозинофилов и отдельные их сегменты обычно отличаются закруглённостью форм; «мостики» и «нити» между сегментами или довольно толсты или почти невидимы, что создаёт в этом последнем случае впечатление разобщённых сегментов.
Моноциты и лимфоциты кроликов не имеют характерных видовых особенностей.
В крови кроликов, по Салин и Вирту, встречаются иногда очень крупные, напоминающие порою эозинофилы, фагоцитирующие клетки — так называемые «клазматоциты». Автор (В. Н.) наблюдал в крови кролика очень большие, фагоцитирующие, с большим количеством вакуолей, эндотелиальные клетки (клазматоциты). Они, однако, никогда не содержали эозинофильных или псевдоэозинофильных гранул. Кровяные пластинки — среднего размера (2,7 µ), довольно многочисленны и имеют более темноокрашенный хромомер.
Морская свинка
Эритроциты диаметром в 4,3—5,7 и до 7,0µ.
В нормальной крови встречается значительное количество полихроматофилов (до 1,0—1,5% от нормо-цитов). Этому соответствует наличие витально гранулированных эритроцитов (0,1—0,9%). У новорождённых морских свинок их количество достигает 20—40%.
Базофилы и эозинофилы крови морских свинок очень напоминают аналогичные клетки крови обезьяны.
Специальные гранулоциты имеют оксифильную, хорошо выраженную, довольно крупную зернистость. Указание Вирта на полное сходство псевдоэозинофилов у кроликов и специальных гранулоцитов у морских свинок следует считать совершенно неправильным: гранулы в псевдоэозинофилах кроликов очень крупны (до 8,5 µ) и окрашены эозином в яркокрасный цвет, специальные же гранулоциты морских свинок имеют гораздо меньшие зёрна блёклой, красновато-розовой, с фиолетовым оттенком, окраски.
Агранулоциты не имеют особенно характерных отличий. Можно только отметить относительную бедность цитоплазмой у больших и средних лимфоцитов.
Для крови морских свинок исключительно характерно наличие у значительной части моноцитов (до 25—40%) так называемых телец Курлова. Это (на хорошо фиксированных препаратах) очень крупные, круглые или овальные, окрашивающиеся азуром в вишнёвый цвет, образования. На менее хорошо фиксированных или долго сохнувших препаратах они как бы сжимаются (возможно, — осмотическое высасывание воды. — В. Н.) и становятся сначала бороздчатыми, а затем приобретают звёздчатую форму. В этом последнем случае на месте, оставленном сжавшимся тельцем, образуется неокрашивающееся пространство (пустота).
Тельца Курлова очень, часто так велики, что оттесняют ядро к периферии, вдавливаются в него и вызывают значительную деформацию не только ядра, но и всей клетки. При очень значительном развитии тельца Курлова могут вызвать распад клетки.
Паппенгейм и Феррата считают, что курловские тельца встречаются не только в моноцитах, но и в больших лимфоцитах. Однако Максимов локализует их только в моноцитах, что представляется более правильным.
Природа и происхождение курловских телец совершенно не ясны. Менее вероятными представляются взгляды на них, то как на вакуоли, наполненные азурофильным секретом, то как на фагоцитированные частички клеток или, наконец, как на стимулирующие рост специальные гигантские гранулы, возникающие под влиянием половых гормонов.
Более вероятны два предположения. По первому, тельца Курлова — это клеточные паразиты из класса простейших. Тогда становится понятным их постепенный рост, угрожающий самому существованию клетки. По другому предположению, курловские тельца — его новообразования в клетках, вызванные внедрением вирусов.
V отдельных взрослых морских свинок и у новорождённых курловские тельца отсутствуют.
Средний размер кровяных пластинок 2—3 µ.
Крыса
Эрптрогшты довольно крупные (5,7—7,0 µ, в среднем 6,2 µ диаметром). Весьма часты полихроматофилы (до 5% у взрослых животных).
Ядро эозинофилов и особенно специальных гранулоцитов развивается по кольчатому типу. Поэтому нередки кольцевидные формы юных и палочкоядерных гранулоцитов.
Эозинофлльные гранулы маленькие, круглые; они густо заполняют цитоплазму.
Зернистость специальных гранулоцитов очень мелкая; она, однако, ясно видна на хорошо фиксированных и окрашенных по Паппенгейму мазках крови.
Белая кровь крыс исключительно лабильна. У двух здоровых крыс одного возраста и даже у одного и того же животного часты значительные колебания в соотношении отдельных видов лейкоцитов.
Мышь
Средний размер эритроцитов 6,7 µ.
Очень часты полихроматофилы (до 10—20% от всех эритроцитов). Иногда встречаются нормобласты.
Белая кровь морфологически весьма близка к крысиной. Только эозинофилы больше размером и имеют довольно крупные зёрна (до 1,0µ).
Моноциты крови мышей, даже при кажущейся норме, часто дают исключительно причудливые, извитые, скрученные ядра, с многочисленными лопастями.
Курица
Картина крови курицы, как и всех птиц, резко отличается от картины крови млекопитающих прежде всего наличием больших эллиптических ядерных эритроцитов. Размеры их часто больше размеров лейкоцитов. Третья группа форменных элементов крови птиц — тромбоциты — тоже имеет ядро.
Это обилие ядерных клеток затрудняет нахождение лейкоцитов в крови птиц, в то время как в крови млекопитающих таких затруднений нет.
Размер эритроцитов курицы 11—13 х 7—8 µ; размер ядер 5—6x3—4 µ. Витально гранулированных эритроцитов довольно много (2—3%, а иногда и значительно больше).
Тромбоциты (веретенообразные клетки) значительно меньше эритроцитов (8,5x5,3 µ).
Лейкоциты птиц, в целом, несколько меньшего размера, чем лейкоциты млекопитающих.
До сих пор нельзя считать решённым вопрос о том, следует ли в крови птиц различать эозинофилов от специальных гранулоцитов (псевдоэозинофилов). Букраба (1928 г.), Ханка (1930 г.), Соловей (1934 г.) и др. склонны признавать существование только одной группы — эозинофилов. При этом В. Букраба (1928 г.), Гедфельд (1911 г.) и Эллерманн и Банг (1908 г.) предлагают диференцировать эозинофилы на круглозернистые и палочкозернистые.
Наоборот, Максимов (1915—1926 гг.), Клинебергер и Карл (1912 и 1928 гг.), Вирт (1925 г.) и школа профессора Н. П. Рухлядева (1930 г.), особенно Л. А. Лебедев (1940 г.), считают, что морфологически и функционально в крови кур различаются псевдо-эозинофилы и эозинофилы. Если, например, псевдоэозинофилы (особенно юные их формы) имеют округлые зёрна, подобные гранулам эозинофилов, то края гранул псевдоэозинофилов кажутся несколько «размытыми», не резкими, по сравнению с гранулами эозинофилов. В подавляющем большинстве псевдоэозинофилов находится палочковидная зернистость, которая часто принимает форму правильных веретён. Ядра зрелых псевдоэозинофилов, так же как и вообще гранулоцитов птиц, более пикнотичны, чем ядра гранулоцитов млекопитающих.
Лебедев (1940 г.) следующими признаками характеризует отличие зернистости псевдоэозинофилов и эозинофилов:
а) Если мазок крови, предварительно окрашенный 0,5-процентным раствором эозина, обработать раствором уксусной кислоты в спирте, то круглые зёрна эозинофилов устойчиво сохраняют красную окраску, а палочковидные и круглые гранулы псевдоэозинофилов обесцвечиваются.
б) Гранулы эозинофилов окрашиваются по Паппенгейму в красно-розовый цвет, а при окраске брилли-анткрезиловой голубой — в голубовато-розовый.
Гранулы же псевдоэозинофилов по первому способу красятся в яркокрасный (иногда коричневато-красный), а по второму — в зеленовато-синий цвет.
При окраске раствором Гимза (даже очень хорошей) зёрна псевдоэозинофилов окрашиваются очень слабо, и поэтому лучшей окраской (и единственно вполне пригодной) для крови птиц следует считать окраску Романовского в модификации Паппенгейма.
в) Оксидазо- и пероксидазо-положительны у кур лишь эозинофильные круглозернистые лейкоциты. Псевдоэозинофилы дают отрицательную реакцию.
г) Суданофильная зернистость есть только у эозинофилов.
д) Ядра у псевдоэозинофилов окрашиваются слабее, чем у эозинофилов.
Вирт считает, что гранулы псевдоэозинофилов даже на юных стадиях их развития не бывают круглыми. Они могут быть продолговатыми, палочковидными, веретенообразными или катушковидными.
По Люндквисту (Lundquist, 1925 г.), гранулы псевдоэозинофилов прижизненно круглые, а различные модификации их форм зависят от фиксации. Прижизненно круглая форма гранул псевдоэозинофилов всё-таки мало вероятна. Однако несомненно, что способ фиксации может несколько влиять на форму зёрен псевдоэозинофилов. Так, автор настоящей книги при некоторых изменениях в способе фиксации получал особую гранулированность псевдоэозинофилов, очень напоминающую мицелий гриба. Близки к этому рн-сунки в работе М. Я. Соловей (1934 г.).
В атласе приведены только те формы клеток, которые принадлежат бесспорно к одной из двух групп оксифильных гранулоцитов кур (и других сельскохозяйственных птиц). Именно, в нём даны эозинофилы, с одной стороны, и палочкозернистые псевдо-эозинофилы, с другой. Практически различить эозинофилы и юные (круглозернистые) формы псевдоэозинофилов крайне трудно.
Лимфоциты кур обычно малые, часто с характерными для них выступами (псевдоподиями), образуемыми цитоплазмой. Последняя в лимфоцитах (даже крупных) скудная.
Моноциты, вопреки Клинебергеру и Карлу (1912—1928 гг.), в крови кур и других сельскохозяйственных птиц, несомненно, есть (школа Н. П. Рухлядева, 1930 г.). Только цитоплазма их серовато-голубая, близкая по окраске к цитоплазме больших лимфоцитов. В цитоплазме моноцитов птиц почти не выявлена мельчайшая азурофильная зернистость.
Очень характерно для крови кур, а также других сельскохозяйственных птиц, наличие на мазках своеобразных фигур распада ядер, — так называемых «теней» ядер, что свидетельствует об очень большой лабильности, «хрупкости» клеток крови птиц. Особенно лабильны клетки очень молодых животных. Это затрудняет подсчёт лейкоцитарной формулы и даже может исказить результаты подсчёта.
Индейка. Гусь. Утка.
Кровь всех этих трёх видов сельскохозяйственных птиц морфологически весьма близка к крови курицы. Несколько вариируют только псевдоэозинофилы. У уток они так же малы, как и у кур; их зернистость не имеет таких отчётливых веретенообразных форм.
Она скорее напоминает неправильной формы короткие палочки, неравной длины. Эозинофилы индеек более крупны, чем псевдоэозинофилы.
У гусей и уток, наоборот, псевдоэозинофилы довольно крупны и значительно превосходят сравнительно маленьких эозинофилов.
Гранулы псевдоэозинофилов очень напоминают зёрна риса. У уток они несколько длиннее и тоньше, чем у гусей.
Тромбоциты крови гуся и утки несколько круглее и короче, чем тромбоциты кур. У индеек тромбоциты, наоборот, более удлинены. Эритроциты имеют несколько более тупые концы.
Клетки крови гусей и уток более резистентны, чем у кур. Поэтому теней ядер в мазках крови гусей и уток гораздо меньше, чем в мазках куриной крови.
Лягушка
Эритроциты очень велики (22,8 х 15,8 µ) и имеют ядра. Количество эритроцитов в крови лягушек невелико (0,38 млн. в 1 мм3).
Базофилы крови лягушки малы, густо наполнены средней величины зёрнами.
Эозинофилы имеют довольно крупные (0,5—1,0 µ), удивительно правильной, круглой формы, раздельно лежащие в цитоплазме зёрна. Цитоплазма интенсивно голубого цвета.
Специальные гранулоциты имеют очень мелкую розовую зернистость. Тип созревания ядра — кольцевой.
Лимфоциты, даже большие, с узкой цитоплазмой, фиксирующейся с вытянутыми псевдоподиями.
В крови нормальной лягушки встречаются моноциты, фагоцитировавшие инородные частички.
Тромбоциты довольно крупные (5x17 µ, чаще несколько короче), эллиптической, вытянутой формы.
Перепечатка материалов без ссылки на наш сайт запрещена