Реферат: Биоиндикация почвы по беспозвоночным
Реферат: Биоиндикация почвы по беспозвоночным
Содержание
Введение
Глава 1. Биоиндикация загрязнения почвы
1.1 История развития и изучения биоиндикации почвы
1.2 Структура животного населения почвы и факторы его разнообразия
Глава 2. Трофическая структура почвенных беспозвоночных
Глава 3. Деятельность беспозвоночных животных в почвообразовании
Глава 4. Материалы и методы проведения исследований
4.1 Деятельность млекопитающих как почвообразующий фактор
4.2 Фаунистическая биоиндикация
4.3 Закономерности зонального распределения комплексов почвенных беспозвоночных
Глава 5. Сообщества мезопедобионтов в критических состояниях
5.1 Влияние техногенного загрязнения на почвенных беспозвоночных
5.2 Влияние других внешних факторов
Выводы
Список литературы
«Человек, в сущности, совершенно не думает о том, что у него под ногами. Всегда мечется… И самое большее – взглянет, как прекрасны облака у него над головой… И ни разу не поглядит себе под ноги, не похвалит: как прекрасна почва!» (Карел Чапек).
Основой жизни на Земле являются круговорот веществ и поток энергии в биосфере. Высокое разнообразие животного мира обусловливает и его разнообразную роль в этих процессах. Связи животных друг с другом, с растениями, микроорганизмами определяют устойчивость биоценозов и экосистем. Животные участвуют в формировании ландшафтов, в почвообразовании, определяют продуктивность различных биогеоценозов и т. д.
Международная система экологического мониторинга, созданная на основе рекомендаций I Международной конференции ООН в Стокгольме в 1972 г. как средство оценки качества окружающей среды и ее изменений (Израэль, 1972; Бурдин, 1985; Криволуцкий, 1994), рассматривает биоразнообразие как один из основных показателей функционирования биоты, в том числе и почвенной. Между тем нелинейная зависимость данных на основе этих показателей порождает трудности введения этого показателя в практику, что определяется несколькими причинами.
В биоиндикации и экотоксикологии почв чаще оценивают структуру населения, биоразнообразие и состояние популяций крупных почвенных беспозвоночных (Гиляров, 1965; Edwards, Bohlen, 1995; Бутовский, 2001), для которых средой обитания является почва как целое. С другой стороны, обитатели почвенных полостей и пор (панцирные клещи, погохвостки, энхитреиды) и обитатели пленок почвенной влаги (нематоды, простейшие) оказываются, в ряде случаев, в большей степени зависимы от действия антропогенных факторов (Криволуцкий, 1983; Hopkin, 1994; Панцирные клещи, 1995; van Straalen, Lokke, 1997; Кузнецова, 2002). Несовпадение реакций разных групп беспозвоночных затрудняет объяснение результатов биоиндикационных исследований (Pokarzhevskii et al., 2003). Среди причин нелинейности ответов популяций одними из основных являются:
1) изменчивость, на уровне «исследуемой точки» (в смысле Мэгарран, 1992), пространственного распределения животных и факторов среды, влияющих на это распределение (Nielsen, 1955; Чернова, Чугунова, 1967; Marinussen, van der Zee, 1996; Ettema, Wardle, 2002);
2) полнота и достоверность учета биоразнообразия (Гиляров, Стриганова, 1975; Edwards, 1995);
3) экологические механизмы отклика популяций на загрязнение (Гиляров, Криволуцкий, 1971; Покаржевский, 1994; Филимонова и др., 2000). Недостаток доступных и сравнимых методов оценки функционирования комплексов почвенных животных в трансформированных почвах (Rombke, Moltmann, 1996) также снижает ценность получаемых биоиндикационных оценок.
В соответствии с этим были выбраны цели и задачи курсовой работы.
Цель работы – изучение особенностей и выявление общих закономерностей структуры и функционирования сообществ мезопедобионтов в естественных и антропогенно изменённых условиях. Задачи исследования:
1. Изучить видовой состав, численность, биомассу, биотопическую приуроченность, трофическую структуру, распределение по почвенному профилю мезонаселения в основных растительных сообществах региона.
2. Выявить роль крупных беспозвоночных в разложении опада и миграции биогенных элементов.
3. Провести трофологические исследования почвенных зоомикробных комплексов. Показать их экобиотехнологические возможности.
4. Выяснить общие принципы структурной и функциональной перестройки сообщества мезопедобионтов, проявления его устойчивости под влиянием природных и антропогенных факторов.
5. Изучить содержание тяжёлых металлов в почвенных беспозвоночных на заповедных и освоенных территориях, проанализировать определяющие факторы. Опробовать аборигенные виды в биоиндикации и экотоксикологическом нормировании загрязнения.
6. Выявить основные факторы организации сообществ и поддержания видовой избыточности мезопедобионтов.
Глава 1. Биоиндикация загрязнения почвы
1.1 История развития и изучения биоиндикации почвы
Интерес к почвенно-зоологическим исследованиям возрос в 40— 50-е годы XX века. Выдающуюся роль в этом сыграли исследования академика М. С. Гилярова и созданной им первой в СССР лаборатории почвенной зоологии (1956), координировавшей исследования в стране. Монография М. С. Гилярова «Особенности почвы как среды обитания и ее значение в эволюции насекомых» (1949), удостоенная Сталинской премии, стала основополагающим трудом в новой области естествознания — почвенной зоологии, вобравшей в себя достижения зоологии, генетического почвоведения, эволюционной теории, а автор — признанным во всем мире основателем почвенной зоологии и ее лидером. В этой книге сформулированы теоретические основы и методологические принципы исследования почвообитающих животных.
В обширном научном наследии М. С. Гилярова, кроме названного труда, выделяются фундаментальные монографии: «Зоологический метод диагностики почв» (1965), удостоенная Государственной премии СССР; «Закономерности приспособлений членистоногих к жизни на суше» (1970), отмеченная премией Московского общества испытателей природы. При непосредственном его участии и под общим руководством вышли «Определитель обитающих в почве личинок насекомых» (1964) и «Определитель обитающих в почве клещей» в 3-х томах (1975—1987), удостоенные Государственных премий СССР.
Почвенная зоология в настоящее время — многоплановая научная дисциплина, изучающая взаимодействие обитающих в почве животных с их средой (почвой) в индивидуальном и историческом развитии.
Почвенно-зоологические исследования охватывают широкий круг современных проблем, связанных с изучением: популяционной структуры и динамики сообществ; специфики биотических связей в сапротрофном зоомикробном комплексе (ЗМК) и структурных особенностей пищевых цепей в почве; роли почвенной биоты в системе биоиндикации антропогенных воздействий на природные экосистемы и биомониторинга окружающей среды. Биоиндикация антропогенных факторов — это определение биологически и экологически значимых антропогенных нагрузок на основе реакций разных организмов и их сообществ (Д. А. Криволуцкий). Почвенная фауна оказывается более чутким индикатором изменений, чем растения, обладающие значительной инерцией по отношению к ним. Использование почвенных беспозвоночных в качестве индикаторных видов оправдано и потому, что наиболее чувствительные к антропогенному воздействию стадии их жизненного цикла проходят в почве: яйца, личинки, куколки.
Биоиндикация — главный метод биологического мониторинга, т. е. мониторинга биоты экосистемы. Но в биомониторинге используются и другие методы. Например, химический анализ содержания загрязняющих веществ в живых организмах.
Почвенная зоология исследует механизмы миграции экотоксикантов в почвах и биологической деградации их. Экотоксикант — токсичное и устойчивое в условиях окружающей среды вещество, способное накапливаться в организмах до опасных уровней концентрации (соединения тяжелых металлов, мышьяка, фтора, углеводородов). Загрязняющие вещества накапливаются в биомассе и мигрируют по пищевым цепям, поэтому в экотоксикологических исследованиях необходимо определять величины биомассы различных групп организмов почвы. В разрушении химических веществ в почве участвуют различные группы организмов: животные, бактерии, грибы, актиномицеты, растения. Они поглощают и перерабатывают химические соединения. В этом блоке исследований выделяются работы казанских педобиологов (Т. И. Артемьева и др.) по изучению роли почвенных беспозвоночных в процессах естественного восстановления биогеоценозов на загрязненных при нефтедобыче территориях и биологической рекультивации. Исследования в производственных условиях дополнены полевыми опытами в почвах лесотундровых, средне- и южно-таежных, лесостепных ландшафтов и в сухих субтропиках. Установлена четкая корреляция естественного восстановления комплекса педобионтов со скоростью распада нефти в почве и техногенной сукцессией растительности. Интенсивность процессов увеличивается с севера на юг: на севере они лимитируются низкими температурами, а в сухих субтропиках — недостатком влаги. Необходимо отметить одно из главных отличий экотоксикологии от классической токсикологии — она исследует реакцию популяции, сообщества и экосистемы на воздействие загрязняющего вещества, а не отдельного организма.
Успешно развивается радиоэкологическое направление, которое прежде всего связано с именем Д. А. Криволуцкого. В биоиндикации радиоактивных загрязнении используется новый для этой области показатель — состояние почвенной биоты. На базе популяционной радиоэкологии можно решать проблемы экологического нормирования и определять факторы биологического риска.
Следует выделить направление, связанное с изучением участия животных в биогенном круговороте химических элементов, их биогеохимической деятельности. Объемный материал по этой проблеме сведен в монографии А. Д. Покаржевского «Геохимическая экология наземных животных» (1985), первой такого плана в почвенной зоологии.
В последние годы приобрели большую актуальность исследования формирования и особенностей структуры экотонных сообществ. Экотоны — переходные пространства между различными природными системами, буферные территории, характеризующиеся постоянно высокой изменчивостью факторов среды. Растущие антропогенные нагрузки увеличивают контрастность и мозаичность экосистем и ландшафтов и, как следствие, формируются новые пограничные экотонные системы и сообщества. В экотонах возникают физические и биогеохимические барьеры для миграции загрязняющих веществ. Почвенные беспозвоночные в таких зонах становятся уязвимыми. В то же время экотоны служат местом сохранения биологического разнообразия.
Среди направлений почвенной экологии, связанных с задачами почвоведения — охрана животного мира почвы. Эффективная система охраны комплексов почвенной фауны возможна при соблюдении двух основных принципов (М. С. Гиляров, Д. А. Криволуцкий, А. Д. Покаржевский). Первый принцип — это охрана экосистем в целом, а не отдельных их компонентов, т. к. контроль каждого вида невозможен. При этом необходимо учитывать, что каждому типу почв соответствует определенный комплекс животных. Второй принцип — это создание системы охраняемых экосистем почв во всех областях страны.
Традиционным, одним из ведущих направлений является изучение почвообразовательной роли почвенных животных. Оно многопланово: изучает влияние отдельных видов на свойства почв (механическое размельчение растительных остатков и вовлечение их вглубь, рассеивание в пространстве экскрементов и т. д.); исследует роль комплексов беспозвоночных в разложении органических остатков и их взаимосвязи с почвенными микроорганизмами в этих процессах. Практический выход имеют работы по использованию отдельных групп беспозвоночных (дождевых червей, диплопод) в зоомелиорации почв. Большое значение придается результатам изучения влияния организационно-хозяйственных мероприятий (распашка земель, орошение, осушение болот, вырубка лесов и др.) на состояние комплексов животных почвы.
Экологический метод диагностики почв, разработанный М. С. Гиляровым, основан на анализе состава животного населения почв, соотношения отдельных его компонентов, численности и экологических особенностей входящих в них популяций. Эти показатели могут быть использованы как индикатор свойств почвы, ее плодородия: каждый вид заселяет те местообитания, где создаются оптимальные условия для его жизнедеятельности. Этот метод с успехом применялся и в тех случаях, когда коррелятивная связь между типом растительности и типом почвы выражена не четко и возникали затруднения в определении типа, разновидности почвы. По степени сходства почвенной фауны сравниваемых участков можно говорить об идентичности типов почв.
1.2 Структура животного населения почвы и факторы его разнообразия
В наземных экосистемах во всех районах Земли обитателями почвы являются 50—99% всех видов животных и на их долю приходится 60—90% наземной зоомассы. Число особей на единицу площади у некоторых групп достигает фантастических величин (до 1 млн. клещей, ногохвосток на 1 кв. м в лесных и луговых почвах). Причины высокого обилия различных представителей почвенного населения изложены в книгах М. С. Гилярова.
Почва представляет очень сложную, многокомпонентную среду. Это трехфазная и полидисперсная система, в которой промежутки между твердыми частицами и их агрегатами заполнены воздухом и водой с растворенными в ней солями. Полидисперсность почвы, или ее гранулометрия, выражается в содержании механических элементов разного размера, от отдельных ионов почвенного раствора до каменистых включений. Разное соотношение фаз создает гамму условий и поэтому для разных размерных групп организмов почва представляет неодинаковую среду, что является одним из факторов высокого разнообразия ее биоты.
В почвенной зоологии принято выделять размерные группы животных, различающихся способами использования среды обитания:
- нанофауна (размеры от микрон до долей мм) – микроскопические объекты, которые могут наблюдаться только с помощью инструментальных методов: простейшие (корненожки, жгутиконосцы, инфузории), мелкие нематоды и коловратки, тихоходки;
- микрофауна (доли мм – доли см) — более крупные немикроскопические организмы: клещи, нематоды, энхитреиды, пауки, коллемболы (ногохвостки), протуры, симфилы, мелкие жуки;
- мезофауна (доли см – несколько см) — крупные беспозвоночные, хорошо различаемые невооруженным глазом, легко учитываемые в полевых условиях при ручной разборке проб почвы: кольчатые и плоские черви, многоножки, пауки, мокрицы, брюхоногие моллюски, насекомые на разных стадиях развития;
- макрофауна (мегафауна) — почвенные позвоночные и крупные формы беспозвоночных.
Установлена обратная зависимость между размерами животных и уровнем их численности (М. С. Гиляров).
Разные размерные группы животных неодинаково используют почву как среду обитания (М. С. Гиляров). Для микроскопических организмов средой обитания оказывается не вся почва, а система микрокапель, капилляров, гравитационной воды, скопления влаги на твердых частицах и между частицами. Когда в почве имеется капиллярная и гравитационная вода, простейшие и коловратки плавают в ней. Животные сохраняют жизнеспособность даже в пленочной воде, находясь в неподвижном состоянии, но не прекращая питания микроорганизмами, детритом, оказавшимися в тех же водяных пленках. Обитатели пленочной воды почв входят также в состав фауны грунтов пресноводных водоемов (Д. А. Криволуцкий). Хотя в почве распределение влаги неравномерное, «прерывистое», но общая поверхность различных форм воды столь велика, что ее можно считать специфическим водоемом, а микроскопических обитателей — физиологически водной экологической группой. Существенное значение для этих организмов имеют особенности почвенной влаги: реакция (рН), химический и газовый состав, наличие почвенных коллоидов, состав водорастворимых солей, особенности органического вещества и порового пространства.
Для мелких членистоногих, называемых микроартроподами, среда обитания — это система ходов и полостей между частицами почвы и их агрегатами, почвенные трещины, ходы более крупных животных и корней, заполненные влажным воздухом. Условия жизни в почве этой группы животных М.С. Гиляров сравнивает с обитанием в насыщенных влагой пещерах. Передвижение обитателей таких пустот не отличается от передвижения по поверхности твердого субстрата. Они могут переживать периоды затопления почвы в отдельных пузырьках воздуха. Все сказанное позволяет считать их физиологически строго наземной экологической группой. Наибольшее значение для этой категории имеют степень порозности и влажности среды, характер распределения органических остатков и гумуса и температурный режим.
Для крупных животных — беспозвоночных и позвоночных — почва представляет среду обитания в целом. Она выступает как рыхлый или плотный и даже твердый субстрат. Движение в плотных слоях почвы встречает большое сопротивление. Передвижение возможно либо по естественным скважинам для животных с тонким змеевидным телом, либо раздвигая частицы почвы, либо копая и прогрызая ходы. При этом животные всегда испытывают механическое воздействие ее твердой фазы и химические воздействия почвенного раствора через покровы, особенно в периоды переувлажнения или затопления почвы. Для крупных почвенных животных имеет значение вся совокупность свойств почвы как единого природного тела.
Почва — это слой наземных биогеоценозов, где происходит разложение, минерализация и гумификация органического вещества. В ней встречаются все стадии разложения животных и растительных остатков: опавшие листья и начинающие гнить листья и корни растений, микроорганизмов. Все это резко расширяет спектр пищевых ресурсов видов и создает возможность одновременного сосуществования в почве животных с различными пищевыми предпочтениями.
Не менее значительным фактором, определяющим видовое богатство почвенной фауны и огромной ее биомассы по сравнению с обитателями других ярусов биогеоценозов, считается то, что животные используют дополнительный резерв белка — из микроорганизмов почвы, а не только от высшей растительности. И здесь не столь важны общие запасы органического вещества, сколько большие ресурсы доступного белка (Д. А. Криволуцкий, А. Д. Покаржевский).
По степени связи с почвой различают три основные группы животных:
- геобионты - проводящие в почве всю жизнь: дождевые черви, некоторые виды многоножек, ногохвосток и др;
- геофилы - у которых какая-то часть цикла развития обязательно проходит в почве: жужелицы, хрущи, комары-долгоножки и др;
- геоксены - случайные обитатели почвы, использующие почву лишь в качестве временного убежища или укрытия: развивающиеся вне почвы пауки, вредная черепашка и др.
Таким образом, широта условий жизни в почве делает ее средой, переходной между водной и наземной. Животные заселили подстилку и минеральные горизонты почвы благодаря специальным адаптациям к различным ее фазам. Расхождение в образе жизни различных размерных групп привело к формированию различных экологических групп — от физиологически водных до строго наземных. Разнообразие источников пищи также обусловило высокий уровень численности, разнообразие видов и экологических групп. Значение почвы в эволюции животного мира заключается в том, что почва рассматривается как среда, через которую животные могли перейти от водного образа жизни к наземному (М. С. Гиляров).
Глава 2. Трофическая структура почвенных беспозвоночных
Комплекс почвенных беспозвоночных включает разные функционально-ценотические группы, различающиеся как по типу питания, так и по форме деятельности. По типу питания выделяются группы:
- фитофаги — животные, питающиеся живыми растительными тканями;
- зоофаги — животные, питающиеся другими животными. К ним относятся хищники и паразиты;
- сапрофаги — животные, питающиеся разлагающимися остатками организмов;
- миксофаги – формы со смешенным питанием.
Наиболее характерной частью почвенного комплекса являются сапрофаги. На их долю приходится основная масса почвенных животных. Биогеоценотическая роль сапрофильного комплекса состоит как в непосредственном биохимическом и физическом возодействии на органические остатки, так и в стимуляции деятельности сапрофитного комплекса (М. С. Гиляров, Ю. И. Чернов). На основе собственного материала и данных мировой литературы Б. Р. Стригановой проведен подробный анализ питания сапрофагов и раскрыты основные особенности детритных пищевых цепей в почве. Комплекс почвообитающих сапрофагов неоднороден по характеру питания входящих в его состав животных. В нем выделяются трофические группировки: фитосапрофаги, микробофаги (микрофитофаги), детритофаги (Б. Р. Стриганова).
Фитосапрофаги утилизируют непосредственно отмершие ткани сосудистых растений. Эти животные способны использовать клетчатку, сапробиотических нематод, гемицеллюлозы, пектины. Наиболее активными разрушителями листового опада являются диплоподы, мокрицы, наземные моллюски, некоторые виды дождевых червей и коллембол, личинки типулид и бибионид. В переваривании грубой растительной пищи у этих форм активную роль играют кишечные симбионты — бактерии, грибы и простейшие — хотя они имеют собственный фермент целлюлазу.
Микробофаги (микрофитофаги) — это потребители бактериальных пленок, микромицетов, почвенных водорослей. К ним относятся многие виды панцирных и тироглифодных клещей, энхитреид, нематод, коллембол, простейших. Животные отличаются наличием специальных энзимов, расщепляющих оболочки и включения грибных клеток. В этой группе имеются потребители почвенных водорослей — альгофаги, как разновидность фитофагов. Но в основном потребление водорослей комбинируется с бактерио- и микофагией. Совместная встречаемость беспозвоночных и водорослевых группировок имеет большое значение в формировании пионерных сообществ в развивающихся почвах (И. В. Стебаев). При выветривании горных пород на рыхлых продуктах их разрушения в первую очередь поселяются водоросли, лишайники и беспозвоночные — альгофаги (микроартроподы). Экскременты животных составляют основу органической фракции первичных почв. Водоросли благодаря водоудерживающей способности создают среду, благоприятную для поселения микроорганизмов (Э. А. Штина, М. М. Голлербах).
Детритофаги потребляют растительные и животные остатки, утратившие исходную структуру и перемешанные с почвой. Эта группа включает дождевых червей, энхитреид, орибатид, личинок некоторых видов жуков и двукрылых, диплопод семейства полизониида, коллембол.
Среди беспозвоночных почвы, потребляющих энергию разлагающихся органических остатков, М. С. Гиляров выделил такие экологические группы, как сапрофаги (детритофаги), микофаги, копрофаги. Копрофаги — питаются собственными экскрементами или других животных (жуки-навозники, личинки серых мясных мух). К сапрофагам в широком смысле слова относят некрофагов — потребителей трупов животных (жуки-мертвоеды, личинки синих и зеленых мясных мух, кожееды). В почве много видов со смешанным типом питания - миксофагов.
Детрит — это измельченные, сильно разложившиеся и гумифицированные остатки животных и растений с населяющими их живыми микроорганизмами, часто перемешанные с частицами почвы. Он представляет последнюю стадию трансформации органического материала. Пищевая цепь, начинающаяся с детрита, называется «детритная пищевая цепь».
Детритная пищевая цепь почвенного яруса экосистемы играет огромную роль в потоке энергии. Согласно многочисленным литературным данным доля потребляемой первичной продукции фитофагами изменяется в зависимости от биоклиматических условий в пределах 2—30% в различных экосистемах. Таким образом, основная масса первичной продукции (98—70%) при отмирании растений поступает в детритный цикл разложения и наряду с продуктами метаболизма и трупами фитофагов и хищников служит источником энергии и элементов минерального питания животным-сапрофагам и микроорганизмам. Большое количество органического материла поставляют в почву и отмирающие корни растений. Схема детритной пищевой цепи на примере экосистемы широколиственного леса приведена на рисунке 1.
Левый вертикальный ряд – это органические остатки в почве на разных стадиях разложения: опад; непереваренные остатки, прошедшие ферментативную и микробиальную обработку в кишечнике наземных и почвенных животных; детрит. В среднем ряду – трофические группировки организмов сапроблока: сапрофитная микрофлора, первичные (фитосапрофаги) и вторичные разрушители (копрофаги относятся к этому блоку).
Особенность почвенной пищевой цепи заключается в том, что продукты биогенной переработки органических остатков одними формами служат пищей для других групп. Хотя многие сапрофаги являются полифагами, но обладают пищевой специализацией в отношении степени предварительного разрушения растительных остатков (механической и химической деструкции органического материала). Фитофаги же, в отличие от сапрофагов, специализированы к биохимическим особенностям видов или групп кормовых растений, у которых прижизненные выделения оказывают аттрактивное или репелентное влияние на животных (Б. Р. Стриганова).
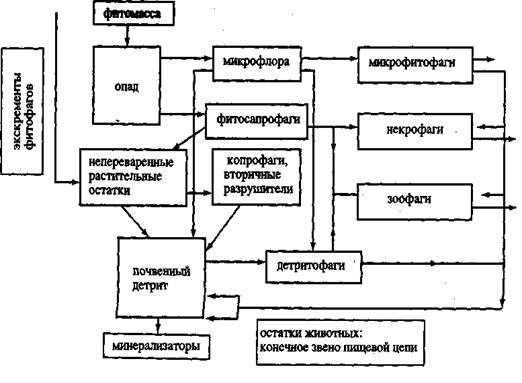
Рисунок 1. Схема детритной пищевой цепи в почве (по Б. Р. Стригановой, 1980)
Среди почвенных сапрофагов по их избирательности к разлагающимся органическим остаткам немецкий ученый В. Дунгер выделил первичных и вторичных разрушителей.
К первичным разрушителям он отнес крупных почвенных и подстилочных беспозвоночных, питающихся отмершей растительностью, но не разложившейся, сохранившей тканевую структуру. Однако растительные остатки начинают перевариваться ими после выщелачивания полифенольных соединений или предварительного разрушения. Животные размельчают и мацерируют частицы растительных тканей в ротовой полости и кишечнике и разрушают связи между отдельными клетками. Они могут быстро размельчать и заглатывать мягкие ткани гниющей древесины, особенно влажной. Совершая вертикальные миграции, беспозвоночные заносят органические остатки в своих кишечниках в глубокие слои и выбрасывают их, формируя копрогенную массу. Этим они стимулируют деятельность сапротрофных микроорганизмов. Существенно то, что первичные разрушители способны переваривать структурные компоненты остатков высших растений — клетчатку, гемицеллюлозы и пектины (Б. Р. Стриганова). Но сапрофитофаги быстрее разрушают «хрупкие» виды листвы нежели остатки с сильно кутикулизированными покровами (корни, кора, листва). К активным разрушителям растительных остатков относятся красные дождевые черви, диплоподы, мокрицы, личинки длинноусых двукрылых (типулиды, бибиониды, ликорииды), некоторые виды коллембол и орибатид.
Доступность и глубина трансформации растительных остатков зависит от механической прочности растительных тканей, содержания трудно разлагающихся веществ — клетчатки, лигнина, пектина и ряда токсичных веществ, угнетающих деятельность микроорганизмов и животных, скорости их выщелачивания (дубильных веществ, танинов и пр.). Так, например, установлено, что буковый опад в широколиственном лесу разлагается очень медленно, слабо потребляется животными. Причиной этого явления оказалось то, что выщелачивание полифенольных соединений из букового опада идет медленно, хотя содержание их не выше, чаи в других породах. В этих условиях первичными разрушителями являются почвенные грибы.
К вторичным разрушителям относятся животные – потребители мертвых растительных остатков, размельченных в кишечнике первичных разрушителей, частично переваренных энзимами животных и микроорганизмов, и обогащенных продуктами их обмена. Они усваивают легкогидролизируемые продукты разложения растительных тканей. Эта категория включает формы со смешанным питанием: сочетание таких пищевых режимов, как сапрофагия, микрофагия, хищничество на мелких животных. Эти беспозвоночные участвуют в процессах деструкции, регулируя состав сапрофитной микрофлоры и способствуя смене грибной фазы разложения микробиальной. Среди почвенных микрофитофагов превалируют потребители микромицетов (Б. Р. Стриганова).
К этой группе относятся копрофаги и детритофаги (дождевые черви, навозники, бронзовики). Значительная доля приходится на мелкие формы (микроартроподы, энхитреиды), обитающие в ферментативном слое подстилки и ризосфере растений, обогащенных разложившимися органическими остатками. Сапрофагия как комплекс пищевых режимов животных показан в таблице 1.
Основной вывод: в комплексе почвенных беспозвоночных - сапрофагов наблюдается последовательный переход к разным пищевым режимам. Наиболее древним пищевым режимом, вероятно, была неизбирательная детритофагия. Черты наибольшей биологической примитивности среди почвенных сапрофагов имеют детритофаги, потребители бактерий, водорослей, простейших. Возможно, на ранних этапах развития почвенного покрова на Земле растительные остатки разрушались микрофлорой.
Таблица 1
Пищевые режимы у беспозвоночных сапрофильного комплекса (по Б. Р. Стригановой)
| Микрофитофаги (микробофаги) | Фитосапрофаги | |
| Первичные разрушители | Вторичные разрушители | |
|
Бактериофагия Микофагия Альгофагия |
Сапроксилофагия | Копрофагия |
| Сапрофиллофагия | Детритофагия | |
| Сапроризофагия | Потребление жидких продуктов разложения | |
Специализация питания почвенных животных шла по пути формирования микофагии и хищничества и, наконец — сапрофагии. Сапрофаги, переваривающие структурные элементы растительных клеток (первичные разрушители), и фитофаги — эволюционно наиболее молодые группы (Б. Р. Стриганова).
Зональная изменчивость комплекса почвообитающих животных имеет ряд тенденций. Общая численность и биомасса их увеличиваются от тундр к широколиственным лесам и луговым степям и снижаются в направлении к аридным областям. В каждой природной зоне складывается свой специфический комплекс животных. Наиболее бедные сапрофильные комплексы характерны для холодных пустынь и тундры, где преобладают микрофитофаги и практически отсутствуют фитосапрофаги. Первичные разрушители появляются в лесотундре и достигают максимума в подзоне широколиственных лесов. В степях обилие и таксономическое разнообразие комплекса сапрофагов уменьшается, но имеются все функциональные группировки. В аридных условиях значение детритных цепей снижается и увеличивается поток энергии через цепи выедания: здесь происходит сочетание сапро- и фитофагии у животных.
Таким образом, особенности субстрата и состав его потребителей определяют специфичность процесса деструкции не только в разных экосистемах, но и в пределах одной экосистемы. Это говорит о том, что для установления реальной картины участия почвенных беспозвоночных в трансформации органических остатков необходимы исследования конкретных экосистем в конкретных условиях.
Глава 3. Деятельность беспозвоночных животных в почвообразовании
Основные функции животных в разложении растительных остатков следующие (Д. А. Криволуцкий):
а) Пропуская через кишечник большую массу растительных тканей, животные размельчают их и тем самым многократно увеличивают суммарную поверхность растительного материала, доступную микроорганизмам, а также для воздействия воздуха и воды.
б) С помощью собственных ферментов и энзимов симбиотических микроорганизмов беспозвоночные расщепляют целлюлозные компоненты клеток и высвобождают лигнин, который находится в сложном соединении с клетчаткой, что имеет большое значение для развития процессов гумификации органических остатков в почве.
в) В ходе пищеварения в кишечнике почвенных беспозвоночных имеет место частичная минерализация растительных остатков, а у некоторых групп — и частичная гумификация. Экскременты животных — одна из составляющих почвенного гумуса.
г) Многие почвенные животные заглатывают вместе с органическими пищевыми веществами минеральные частицы почвы, способствующие перетиранию в кишечнике пищи. Минеральные частицы (глинистые, песчаные) перемешиваются в кишечниках, спрессовываются и склеиваются выделениями кишечника, образуя разной величины «структурные отдельности» почвы — зернистые комочки; чем выше их количество, тем плодороднее почва.
д) Совершая вертикальные миграции в почве, животные заносят растительные остатки в глубокие горизонты и перемешивают органические и минеральные частицы. Передвижения животных способствуют улучшению условий аэрации почвы, что, в свою очередь, стимулирует аэробные процессы разложения органических остатков.
Исследования круговорота элементов в наземных сообществах показывают, что большое значение в питании беспозвоночных имеет азот и его соединения (А. Д. Покаржевский, Д. А. Криволуцкий). Но внесение азотных минеральных удобрений не оказывает положительного эффекта на численность животных. Это связано с тем, что животные используют в пищу не минеральные формы азота, а органические его соединения — белки, витамины. Питательная ценность белков зависит от их аминокислотного состава. Животные не способны синтезировать незаменимые аминокислоты: лизин, гистидин, аргинин, треонин, валин, метионин, изолейцин, лейцин, фенилаланин, триптофан. Их достаточно только в животном белке и в белках микроорганизмов. Незначительное количество аминокислот в почве и растительных остатках свидетельствует о том, что сапрофаги могут испытывать белковый голод. Существуют два пути преодоления дефицита аминокислот: хищничество и использование микробиальной массы экосистемы. Дефицит белка восполняется и копрофагией. Известны случаи «автокопрофагии» у мокриц и диплопод (Б. Р. Стриганова): животные потребляют растительный опад, который переваривают в очень небольшой степени, а также собственные экскременты, полежавшие в почве и заселенные микрофлорой, которая активно разрушает лигно-целлюлозный комплекс. В экскрементах животные усваивают легкоусвояемые продукты микробного распада структурных компонентов растительных тканей – это основной энергоресурс. Недостаток белковой пищи удовлетворяется за счет микробной массы. Автокопрофагия характерна для животных, у которых отсутствует комплекс облигатных симбионтов в кишечнике.
Почвенные беспозвоночные имеют симбиотические отношения с разными группами микрофлоры — аммонификаторами, фиксаторами молекулярного азота и разрушителями клетчатки. Перерабатывая растительные остатки в пищеварительном тракте животного, они обеспечивают его необходимыми аминокислотами. Но в пищеварительной системе может развиваться и не симбиотическая микрофлора, поступающая с пищей. Очевидно, поглощение почвы тоже связано с потреблением микроорганизмов.
Азот и фосфор для различных групп беспозвоночных — лимитирующие элементы. Азот в виде белковых тел является конструктивным материалом живых организмов биосферы, фосфор в виде молекул органических фосфатов является преобразователем энергии в биосфере. Связь между этими элементами реализуется благодаря деятельности микроорганизмов в экосистемах суши.
С деятельностью различных групп животных почвоведы связывают образование различных форм гумуса (W. Kubiena A. Brauns и др.).
Грубый гумус-мор. Отличается кислой реакцией, малым количеством экскрементов животных. Содержит неразложившиеся и слабо разложившиеся грубые растительные остатки, пронизанные гифами грибов. Отсутствуют прочные связи органических веществ с минеральной глинистой составляющей. Такой гумус образуется во влажном и прохладном климате, когда опад богат трудно разлагаемым материалом — лигнином и целлюлозой, как под хвойными породами. В формировании такого перегноя участвуют раковинные амебы, клещи-орибатиды, ногохвостки. Участие животных незначительно, преобладает грибной тип разложения. Мор свойственен преимущественно подзолистым и дерново-подзолистым почвам.
Мягкий гумус-мулль. В его структуре заметны лишь мелкие неразложившиеся растительные фрагменты. Это результат интенсивной деятельности энхитреид, дождевых червей, многоножек, мокриц, личинок насекомых. В кишечнике животных органическое вещество растительных остатков претерпевает сложные превращения и перемешивается с минеральными частицами. В результате образуется органо-минеральный комплекс. Особенно заметна роль крупных дождевых червей, которые, вовлекая растительные остатки с поверхности в глубь почвы и перемешивая их с минеральными частицами почвы, способствуют созданию зернистой структуры и углублению аккумулятивного горизонта. Кроме того, накапливаются водопрочные экскременты разных животных. Эта форма гумуса присуща серым и коричневым лесным почвам под лиственными лесами и черноземам.
Промежуточная форма гумуса-модер (волокнистый гумус) имеет умеренно кислую реакцию. Он образуется благодаря жизнедеятельности мелких членистоногих, экскременты которых смешиваются с минеральными частицами. В нем умеренное содержание разложившихся фрагментов растительности: структура измельченных частей еще различима, и они не пронизаны гифами грибов. Лучше всего эта зоогенная форма гумуса образуется в условиях повышенной влажности, которая складывается в нижних влажных слоях подстилки под более свежим опадом. В этих условиях в переработке подстилки принимают участие и крупные беспозвоночные.
Но нужно иметь в виду, что в одинаковых растительных сообществах могут формироваться различные типы гумуса. Это зависит от различий в характере почвенной фауны, микрофлоры, абиотических факторов. Так, в буковых лесах на северо-западном Кавказе, где очень бедное почвенное население, отсутствуют черви, подстилка медленно разлагается, слеживается в виде плотного толстого пласта, прочно переплетенного грибным мицелием. Образуется грубый перегной. А в буковых лесах Закарпатья почва богата членистоногими и червями, подстилка быстро разлагается, образуется мулль (Г. Ф. Курчева).
Взаимосвязь компонентов органического комплекса почвы: корневые системы — беспозвоночные — гумус установлена М. С. Гиляровым (1965). Содержание и распределение гумуса по почвенному профилю коррелирует с вертикальной миграцией различных видов беспозвоночных, их численностью, биомассой, пищевой избирательностью и активностью.
Рассмотрим почвообразовательную деятельность отдельных групп беспозвоночных, населяющих разные типы почв, в т. ч. и юга России.
Дождевые черви. Большинство дождевых червей относится к семейству люмбрицид (Lumbricidae). Черви встречаются в разных природных зонах. Они отсутствуют в полярных пустынях, а в полупустынях и пустынях, в неокультуренных зональных почвах. Могут обитать лишь в оазисах или приречных незаселенных почвах. Наибольшей плотностью и наибольшим видовым разнообразием люмбрицид характеризуются зоны смешанных и широколиственных лесов, лесостепь и интразональные луговые ценозы. К югу от лесостепи зоомасса дождевых червей начинает снижаться. В черноземных почвах степей распространены только собственнопочвенные формы дождевых червей и то только в почвах плакоров в очень небольшом числе и только в западных районах степной зоны. Но большая численность может быть в интразональных биотопах (в почве лугов, речных террас, на склонах оврагов) и вблизи жилья, в антропогенных биотопах (на огородах, в компостах и т. п.).
По характеру используемой пищи среди дождевых червей выделяют два морфо-экологических типа. Первый тип — виды, питающиеся на поверхности почвы слабо разложившимися растительными остатками и даже зелеными частями травянистых растений (первичные разрушители по Дунгеру). Например, Lumbricus terresticus. Второй тип — собственнопочвенные черви, питающиеся перегноем почвы. Например, Nicodrilus caliginosus. Представители второго типа значительно энергичнее перемешивают и рыхлят почву, чем первого.
По характеру вертикального распределения по профилю почвы поверхностнообитающие формы (первый тип) делят на три группы: подстилочные — в деятельном состоянии населяющие подстилку и не мигрирующие даже при пересыхании или промерзании почвы глубже 5—10 см; почвенно-подстилочные черви, мигрирующие глубже 10—20 см лишь при неблагоприятных сезонных условиях; норниковые — имеющие постоянно глубоко уходящие (на 1 м и более) ходы, которые они обычно не покидают. Собственнопочвенные дождевые черви (второй тип) не выходят на поверхность почвы при питании или спаривании. Они могут быть разделены на две группы: роющих — обитателей глубоких горизонтов, откуда они мигрируют вглубь только при неблагоприятных гидротермических условиях; норников — имеющих постоянные глубокие ходы, но питающихся также в гумусовом горизонте почвы.
Дождевые черви способствуют минерализации и гумификации растительных остатков, стимулируют деятельность микрофлоры. У них большая потребность в азоте. Предполагается, что ограниченность запасов азота в минеральных горизонтах почвы стала причиной перехода червей к питанию мало разрушенными растительными остатками. Благодаря прохождению пищи через кишечник червей и усилению развития микрофлоры в их экскрементах почва обогащается фульво- и гуминовыми кислотами. Они повышают подвижность азота, калия, фосфора, магния, кальция. При высокой численности содействуют нейтрализации кислых почв: их известковые железы выделяют кальций в кишечный тракт, где воздействуют на пищу. Дождевые черви обогащают почву витаминами группы В, РР, белками: они содержатся в организме животных и после гибели их выделяются в окружающую среду.
Благодаря размножению микрофлоры в кишечнике червей происходит ускорение разложения органики по сравнению с почвой. В экскрементах червей ускоряется минерализация органики: там резко возрастает численность актиномицетов и других микроорганизмов. Дождевые черви способствуют развитию простейших (жгутиконосцев, инфузорий, саркодовых) в почве и увеличению их разнообразия. В целом дождевые черви положительно влияют на биологическую активность почв.
Несомненный интерес представляют диплоподы (Diplopoda), или двупарноногие многоножки. Они играют большую роль в процессах первичного разложения и минерализации органических остатков. В подавляющем большинстве это фитосапрофаги, т. е. питаются опавшей листвой, ветками, гниющей древесиной, предварительно размельченными в ротовой полости. Высокая усвояемость пищи у диплопод объясняется высокой активностью целлюлазы в кишечнике. Собственная целлюлазная активность усиливается ферментами симбиотических микроорганизмов. В кишечнике происходит мацерация растительных тканей. Кроме того, многие виды регулярно потребляют собственные экскременты. При дефиците влаги охотно переходят к фитофагии. Питание живыми растениями восполняет дефицит влаги в организме. Но в силу этого в агроценозах диплоподы становятся вредителями сельскохозяйственных культур. Высокая пищевая активность, способность к потреблению слабо разрушенного опада, высокая усвояемость пищи делают их важнейшим агентом почвообразования (С. И. Головач).
Очень важна роль диплопод в круговороте кальция (Ю. Б. Бызова). Они накапливают кальций в скелете и теле. Получая кальций с пищей, диплоподы в течение жизни выделяют его в виде простых соединений с экскрементами, личиночными шкурками и при отмирании, обогащая им поверхностный слой почвы. Диплоподы могут быть индикаторами почв с высоким содержанием кальция.
Диплоподы широко распространены в различных природных зонах. Наиболее предпочитаемым местом обитания является лесная подстилка широколиственных лесов. Их встречаемость и обилие уменьшаются в почвах открытых ландшафтов — лугов, степей, полупустынь, пустынь и обработанных земель. Они отсутствуют в зоне тундр.
Диплоподы обитают в опаде и верхних горизонтах почвы, уходя в периоды покоя на глубину до 30 см или в гнилую древесину. Они чувствительны как к недостатку, так и избытку влаги в среде: не употребляют излишне увлажненную, с капельками воды, пищу. Нормально двигаются и питаются при хорошей аэрации подстилки, поэтому избегают уплотненных нижних слоев мощной подстилки, где идут гнилостные процессы.
Весьма обычны диплоподы в степной зоне европейской части страны. Кивсяк— характерный элемент сапрофильного комплекса почвенных беспозвоночных в различных биогеоценозах степной зоны Нижнего Дона. Мозаичность почвенно-растительного покрова и, соответственно, гидротермического режима обусловливают пестроту пространственной структуры вида. Кивсяки заселяют байрачные и пойменные леса, овражно-балочные лесонасаждения, лесополосы, заросли кустарников и степные участки. На юго-востоке Ростовской области в полупустынных ландшафтах этот вид отсутствует. Контрастность показателей численности и биомассы свидетельствует, что он предпочитает лесонасаждения и естественные леса. Большое значение играет природный состав насаждений: кивсяки проявляют пищевую избирательность к различным видам лесного опада и ветоши. Это определяет темпы разложения растительных остатков и характер почвообразовательных процессов. Этот вид кивсяка может быть использован для оптимизации процесса разложения в условиях изменяющейся экологической обстановки в разных ландшафтах юга России.
Жуки-чернотелки — характерный и массовый элемент аридных биогеоценозов. Это организмы — минерализаторы, которые производят глубокую переработку опада, препятствуя его трансформации в гумус, сдвигая его в сторону минерализации. Кроме непосредственного поедания подстилки, чернотелки разлагают подстилку, измельчая ее и увеличивая тем самым поверхность разлагающегося субстрата. Экскременты чернотелок — центры стимуляции целлюлозоразрушающей микрофлоры, что дополнительно усиливает разложение подстилки. В ряду зональных степных биогеоценозов уровни численности и биомассы чернотелок увеличиваются от северного предела степей к сухим, после чего снова уменьшаются.
Глава 4. Материалы и методы проведения исследований
4.1 Деятельность млекопитающих как почвообразующий фактор
Изменения в экосистемах, вызываемые позвоночными животными, накапливаются и в конечном итоге накладывают отпечаток на весь облик территории и характер почвообразования. Непосредственно на почвенный покров воздействует роющая деятельность млекопитающих. Подавляющее большинство исследований роющей деятельности выполнено в открытых ландшафтах (степи, полупустыни, пустыни), где отсутствие готовых естественных убежищ является причиной активной деятельности млекопитающих по сооружению нор в почве (Б. Д. Абатуров). По свидетельству А. Н. Формозова только в степной зоне Евразии из 92 степных млекопитающих жизнь 72 видов связана с норами. Наиболее типичными представителями степных млекопитающих являются сурки, цокоры, суслики, степная пищуха, джунгарский хомячок, тушканчики, полевки.
Еще в 1906 году профессор Н. А. Димо и ботаник Б. А. Келлер изучали деятельность малого суслика на юге Саратовской губернии. Их совместная работа содержит массу интересных фактов. Они установили, что не только химический состав, но и структура почвы в суслиных холмиках отличаются от почвы, не затронутой роющей деятельностью мелких млекопитающих. Благодаря своей рыхлости, пористости и трещиноватости эти холмики весной легко насыщаются влагой, поскольку в зимнее время снег собирается вокруг них в большем количестве, чем на ровной поверхности степи. Поэтому весной растительность на них развивается с особенной энергией. Испарение поглощенной влаги из рыхлых холмиков происходит медленнее, чем из плотной почвы.
На основе накопленных сведений о роющей деятельности позвоночных животных выделен ряд форм их воздействия на среду (Б.Д. Абатуров).
а) Норы животных разрыхляют почву, улучшают ее аэрацию, способствуют более глубокому увлажнению почвенной толщи водами атмосферных осадков, защищают почвенную влагу от непродуктивного физического испарения.
б) При рытье нор животные выносят на поверхность материал глубоких горизонтов почвы и тем самым увеличивают содержание легкорастворимых солей, гипса и карбонатов в верхних слоях почв.
в) На перерытых участках вследствие более интенсивного увлажнения происходит проседание почвенной толщи, формируются отрицательные формы микрорельефа.
г) Накопление почвенного материала, вынесенного на поверхность при рытье нор, вызывает формирование положительных форм микро-и нанорельефа (кучки и холмики выброшенной земли)| с иными физико-химическими свойствами почв.
д) В местах постоянного расположения нор животные обогащают почву химическими веществами за счет экскрементов и тем самым меняют ее химический состав и улучшают плодородие.
е) Роющие животные перемешивают верхний гумусовый горизонт с нижележащей материнской породой и тем самым увеличивают мощность этого горизонта.
ж) Перерытый норами и разрыхленный почвенный материал легко поддается действию ветра и воды, что служит причиной размывания и развеивания почв и образования эрозионных форм рельефа.
з) Перерытые и нарушенные роющей деятельностью участки заселяются сорными и полевыми видами растений и почвенных беспозвоночных и тем самым служат причиной формирования специфической сорно-полевой фауны и флоры.
и) В результате роющей деятельности меняется не только состав растений, но и их масса, при этом изменения могут быть направлены в сторону как уменьшения (при засыпании растений выброшенной землей, обеднении почвенного субстрата), так и в сторону увеличения (при улучшении плодородия почвы).
к) Разрастание на перерытых местах сорных видов растений вызывает формирование залежного растительного покрова. Локальное нарушение свойств почвенно-растительного покрова служит причиной микрокомплексности почв и растительности в степных, полупустынных и пустынных ландшафтах.
Эти положения, сформулированные еще несколько десятилетий назад (А. Н. Формозов; А. Г. Воронов; В. В. Кучерук и др.), остались без существенных изменений по настоящее время.
В девяностые годы прошлого века обострилась проблема влияния роющих мелких млекопитающих на эрозию почв в горных регионах. X. П. Мартинес Рика, М. П. Пар до Ара описали влияние обыкновенной полевки на протекание эрозионных процессов на южном макросклоне Центральных Пиренеев. Норы обыкновенной полевки представляют собой хорошие водоводы как в период снеготаяния, так и во время сильных или затяжных дождей. Весной легко заметны потоки воды, устремляющиеся по системе ходов и нор. В том случае, если потоки воды не канализированы, почва насыщается влагой, а ее избыток стекает по поверхности, где растительный покров задерживает частички почвы, увлекаемые потоком. В то же время в местах, населенных полевками, возникают другие условия, при которых водный сток приобретает важную эрозиогенную функцию. Если норы протяженны и неглубоки, располагаются вблизи небольших естественных русел, они могут перехватывать водные потоки и топографически замещать эти русла. При такой ситуации формируется новое русло стока, порождаемое сетью заброшенных нор полевок. В местах с известняковым субстратом, где распространены карстовые явления, мелкие млекопитающие играют важную роль в подземном стоке. При возникновении расщелин на горных пастбищах, прорезающих почву, начинается процесс оврагообразования.
Косвенное воздействие роющей деятельности грызунов имеет большее значение, чем прямое отторжение части почвенного покрова.
Заметные изменения в структуре природных комплексов вызывает выпас скота в степи. Растительный покров разреживается, понижается проективное покрытие, уменьшается высота растений. В процессе выпаса травоядные копытные животные поедают молодые растения, не давая достигнуть им полного развития, начинают преобладать растения, способные к быстрому отрастанию и вегетативному возобновлению. Чрезмерный выпас приводит к уменьшению числа многолетних растений и увеличению однолетних. В пастбищном травостое появляются непоедаемые растения: ядовитые, жесткие, опушенные, колючие.
Юго-восточная часть Ростовской области (Заветинский район) входит в зону сухих и отчасти полупустынных степей, сформированных на каштановых и солонцовых почвах. Территория используется под пастбище. Интенсивный, бессистемный выпас скота привел к усиленному уничтожению и вытаптыванию растительности, исчезновению ветоши, разрыхлению верхнего горизонта почвы, которая в силу этого подвергается эрозии. Высокая степень дигрессии пастбищ явилась причиной обеднения экологических ниш животных, что отрицательно сказалось на всем их комплексе. Наши исследования подтвердили имеющиеся в литературе сведения (М. С. Гиляров, Т. С. Перель) о том, что в каштановых почвах практически исчезают дождевые черви, личинки длинноусых двукрылых, сильно уменьшается количество энхитреид, мокриц, кивсяков — активных почвообразователей. Ни в одном из подтипов каштановых почв нами не обнаружены дождевые черви. Деятельность беспозвоночных в глубоких горизонтах часто лимитируется в зоне каштановых почв их засолением (М. С. Гиляров). Большую устойчивость к такому стрессовому воздействию выработали личинки хрущей, долгоносиков, мух. Слабая сформированность комплексов педобионтов — причина заторможенности деструкционных процессов.
4.2 Фаунистическая биоиндикация
Одним из важных показателей антропогенного воздействия на окружающую среду является изменение видового состава и количества почвенных и напочвенных беспозвоночных животных и метод лихеноиндикации.
Для определения антропогенного воздействия на экосистему в различных точках урболандшафта применялся метод фаунистической биоиндикации. Дождевые черви, обитающие в верхних слоях почвы, подвергаются воздействию токсикантов и характеризуют первые стадии загрязнения почв, они очень четко реагируют на природные и антропогенные изменения.
Для этого отбираются пробы почвы для количественного учета дождевых червей в нескольких исследуемых точках. На площадках в каждой точке размером 10х10 метров берутся пробы с трех площадок размером 25х25см. с глубины 20 см.
Процесс взятия пробы проходит следующим образом. Сначала отмечают площадь пробы, забивая по углам квадрата колышки, натягивая между ними шнур. Рядом с площадкой раскладывают клеёнку, на которую потом помещают выбираемую лопатой из пробы почву. Вынутые на клеенку небольшие порции почвы, тщательно перебирают руками. Распределяют по поверхности клеёнки тонким слоем. Затем длинным пинцетом собирают дождевых червей, пересчитывают, а затем отпускают. Данные записываются в протокол отбора проб с каждой площадки. При изучении дорожно - тропиночной сети и других форм антропогенного воздействия на почву подсчитывают численность дождевых червей по уровням удаления от объекта загрязнения.
Выполнение этих исследований позволяет:
Охарактеризовать загрязнение почвы и его вредное воздействие на экосистему.
Установить влияние загрязнения почвы на численность дождевых червей.
Установить степень загрязнения почвы в обследуемой местности.
Своевременно обнаружить загрязненность почвы по количественному учету дождевых червей и вести контроль, за состоянием почвы в условиях антропогенного воздействия.
4.3 Закономерности зонального распределения комплексов почвенных беспозвоночных
Важнейшей характеристикой структуры животного населения почв является соотношение трофических групп. На основе обобщения материалов разных авторов установлены общие закономерности зональных изменений почвенного комплекса животных (М. С. Гиляров, Ю. И. Чернов). От гумидных районов в к аридным снижается доля сапрофагов и возрастает роль фитофагов. От тундр до широколиственных лесов в животном населении зональных почв господствуют сапрофаги, составляя 80—99% общей зоомассы. Во всей лесной зоне основу комплекса сапрофагов составляют дождевые черви, энхитреиды, частично нематоды. В тундровой зоне равноценную с этими животными роль выполняют типулиды (двукрылые). Наиболее разнообразен комплекс сапрофагов в широколиственных лесах и в лесостепи: кроме дождевых червей и энхитреид, здесь существенную роль в почвообразовании выполняют кивсяки и мокрицы. С увеличением аридности условий к югу от лесостепи резко снижается доля сапрофагов в животном населении и резко увеличивается доля растительноядных. Сапрофаги составляют не более трети всей зоомассы (Ю. И. Чернов). Важно отметить, что значительная часть фитофагов заселяет почвенный ярус. Для таких переходных условий особенно характерно обилие факультативных почвенных фитофагов: личинок хрущей, чернотелок, некоторых мертвоедов, щелкунов, пыльцеедов и т. д.
В плакорных сообществах полосы сухих степей, пустынь и полупустынь почти полностью исчезают типичные сапрофаги, все фазы развития которых связаны с почвой, а сохраняются некоторые личиночные формы, непродолжительно пребывающие в почве или связанные с особыми локальными условиями.
В аридных зонах растительноядные позвоночные животные как по массе, так и по функциональной значимости в экосистеме не уступают беспозвоночным (Ю. И. Чернов; Ю. И. Чернов, К. С. Ходашева, Р. И. Злотин).
Аналогичная картина наблюдается в горных условиях. Так, сопоставление различных типов сообществ в горных районах Внутреннего Тянь-Шаня (Р. И. Злотин) отчетливо выявило усиление дифференциации трофического состава животного населения. В системе высотных поясов по мере изменения типа сообществ (влажные луга, остепненные луга, холодные степи, сухие степи, полупустыни, пустыни) увеличивается удельный вес фитофагов и хищников по сравнению с сапрофагами, но общая зоомасса падает.
В целом по мере увеличения суровости гидротермических условий существования в почве животные, участвующие в разложении, оказываются в более тесной связи с надземным блоком живой зеленой массы (И. В. Стебаев). В степях и пустынях среди беспозвоночных-деструкторов наиболее активной группой оказываются саранчовые. Поедая большую массу зеленых растений, они слабо ее усваивают, что приводит к накоплению экскрементов, масса которых во много раз превышает массу их тел. В результате разрушения челюстями клеточных оболочек растений внутри кишечника создаются условия для бурного развития микроорганизмов — типичных представителей почвенной микрофлоры, попадающей в кишечник с пищей (аммонификаторов, целлюлозоразрушителей и др.) (И. В. Стебаев, Н. Н. Наплекова и др.). Интенсивное разложение растительной массы совокупным действием ферментов и микроорганизмов насекомого увеличивает запас питательных веществ, высвободившихся из растительной массы. Часть микроорганизмов со временем отмирает, лизируется и становится источником белковой пищи фитофагов, другая часть — выбрасывается с экскрементами. Значение этого процесса особенно велико: зеленая масса сразу переходит в стадию разлагающейся подстилки, минуя длительные стадии ветоши и войлока.
Таким путем насекомые-фитофаги надземного яруса биогеоценоза ускоряют нисходящую ветвь круговорота веществ. Роль саранчовых очень важна для районов, характеризующихся дефицитом влаги в почве, потому что они, с одной стороны, значительно раздвигают пространственные границы процессов почвообразования выше поверхности самой почвы, а с другой — значительно ускоряют эти процессы (И. В. Стебаев, Л. Б. Пшеницына, О. Н. Шиляева).
Таким образом, пищевые связи животных и трофическая структура их комплексов являются ведущими факторами при определении их роли в почвообразовательных процессах.
Глава 5. Сообщества мезопедобионтов в критических состояниях
5.1 Влияние техногенного загрязнения на почвенных беспозвоночных
Проведение модельного полевого опыта по влиянию сернистого ангидрида на педобионтов позволило получить фактические данные о структурных изменениях и проявлениях устойчивости сообщества при однократном «ударном» и слабых краткосрочных воздействиях.
Под воздействием комплекса отходов промышленных предприятий (источник эмиссии - ИЭ) отмечены все выявленные в полевых экспериментах признаки нарушения стабильности сообщества. Общая численность мезопедобионтов в непосредственной близости от ИЭ снижается почти вдвое, биомасса — более чем в пять раз относительно контроля. Это происходит за счет значительного сокращения численности литобиид, геофилид, диплопод, личинок двукрылых. При этом вблизи ИЭ фиксируются наиболее широкие флуктуации плотности большинства групп беспозвоночных во времени, что является признаком глубоких, долгосрочных, а возможно, и необратимых функциональных изменений сообщества.
Изменяется качественно и видовой состав ряда доминирующих групп. В импактной зоне исчезают некоторые моллюски, геофилиды, диплоподы, личинки двукрылых, преобладающие на фоновой территории. В то же время вблизи ИЭ появляются дождевые черви, повышается обилие энхитреид и личинок долгоносиков. Эти факты свидетельствуют о разной реакции беспозвоночных на изменения среды обитания под воздействием отходов сернокислотного производства.
|
|
|
|
|
|
|
|
|
|
|
|
|
|

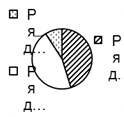
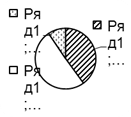
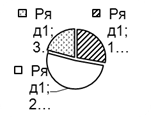
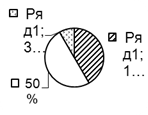
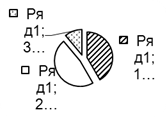
Рис. 2. Соотношение плотности трофических групп мезопедобионтов обследованных участков лиственничника в течение трёх лет (I, II, III) по мере удаления от источника эмиссии (У 1 → У 3): 1 – хищники; 2 –сапрофаги; 3 –фитофаги.
Таблица 2
Изменения видового состава некоторых групп беспозвоночных (экз/м2) лиственничника под воздействием отходов промышленных предприятий
| Виды | Расстояние от ИЭ, км | ||
| 0,8 | 2 | 4,5 | |
|
Gastropoda Discus pauper Euconulus fulvus Nesovitrea hammonis |
- - - |
15 2 1 |
17 1 2 |
|
Diplopoda Underwoodia kurtschevae Pacifiiulus amurensis Angarozonium amurense Uniramidesmus dentatus |
8 - - - |
3 - 2 0,3 |
6 4 21 - |
|
Geophilomorpha Arctogeophilus macrocephalus Geophilus sp. Escarius japonicus Strigamia sulcata |
11 1 - - |
50 12 8 1 |
47 17 12 6 |
Внутри сообщества мезопедобионтов вследствие промзагрязнения существенно возрастает пресс хищников (рис. 5). Сильнее других откликаются сапрофаги, при этом в одних случаях речь может идти об ингибировании, а в других – об активизации их деятельности. Это определяется составом поллютантов, способных либо подкислять, либо подщелачивать почву. Появление земляного червя Eisenia nordenskioldi и исчезновение многоножек A. amurense и E. japonicus вслед за изменениями кислотности обитаемого слоя есть тому подтверждение.
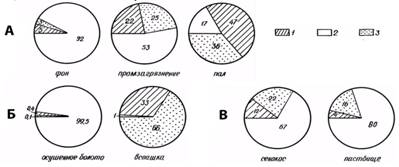
Рис. 3. Соотношение трофических групп почвенной мезофауны (% от общей биомассы): 1 – хищники, 2 – сапрофаги, 3 – фитофаги; А — лес, Б — болото, В — луг.
Кроме того, беспозвоночные одной трофической группы неоднозначно откликаются на одно и то же воздействие, что неоднократно отмечалось и другими исследователями. Среди крупных сапрофагов численность минерализаторов (моллюски, диплоподы) снижается более чем в тридцать раз, а гумификаторов (энхитреиды, люмбрициды), напротив, возрастает. В результате соотношение функциональных трофических группировок изменяется в 40 раз.
Для характеристики влияния отходов производства на видовую структуру комплекса сапрофагов были рассчитаны выровненность по Пиелу и информационный индекс Шеннона. По мере приближения к ИЭ эти показатели уменьшаются с 1,85 до 0,16 и с 2,16 до 0,28 соответственно. Это свидетельствует об упрощении структуры сапротрофного блока, росте численности отдельных видов.
5.2 Влияние других внешних факторов
Аналогичная картина изменения пространственной структуры и динамики сообщества прослеживается при пожарах и рубках леса. При этом типе воздействия также отмечается резкое снижение общей биомассы педобионтов при нарастании степени нарушенности биотопа. Аналогично реагирует на это и их трофическая структура. Пирогенные перестройки сообщества мезопедобионтов вызваны, в первую очередь, разрушением среды обитания, а именно физического компонента экологической ниши. Происходит это в основном за счет полного исчезновения или уменьшения мощности подстилки, что сводит к минимуму объём обитаемого пространства, следовательно, вызывает рост конкуренции на участках, слабо затронутых огнём (рефугиумах). При уменьшении абсолютной численности беспозвоночных в основном угнетаются обитатели подстилочного горизонта. Пагубное влияние ежегодных палов на луговых педобионтов, особенно поверхностно обитающих, отмечается неоднократно. Такой характер изменения вертикальной структуры мезопедобионтов является закономерным для множества подобного рода воздействий.
На открытых местообитаниях периодические затопления также значительно обедняют комплексы почвообитающих животных. На лугах очень мало хищников, из них полностью отсутствуют подстилочно-почвенные литобииды и геофилиды. Из сапрофагов не встречаются подстилочные диплоподы, личинки бибионид и типулид. Эта трофическая группа представлена фактически одним видом земляных червей.
Механизированные ежегодные сенокосы монокультуры угнетающе влияют на биомассу мезопедобионтов. Напротив, умеренный выпас благоприятен для почвенных обитателей, в то время как чрезмерное выедание и вытаптывание пагубно сказываются на поверхностно-обитающих педобионтах (рис. 4).
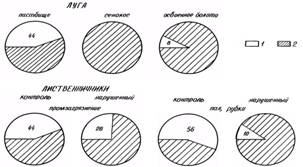
Рис. 4. Соотношение численности (% от общей) поверхностно-подстилочных и собственно-почвенных обитателей: 1-поверхностно-подстилочные, 2 - собственно-почвенные.
При анализе влияния различных возмущающих факторов на характер структурных изменений сообщества мезопедобионтов становится очевидной неспецифичность внешнего воздействия, т. е. эффект от влияния не связан с происхождением возмущения. Аэрополлютанты, пожары, промышленные рубки, многолетняя вспашка осушенных болотных массивов одинаково влияют на беспозвоночных, вызывая:
1. Снижение численности/биомассы;
2. Депрессию и гибель сапрофагов, и в первую очередь, минерализаторов;
3. Усиление пресса хищников;
4. Смену типа трофической цепи с детритной на пастбищную;
5. Деградацию подстилочного комплекса;
6. Изменения пространственного распределения;
7. Обеднение видового состава.
Чрезмерный сенокос на плакоре также негативно сказывается на почвенных беспозвоночных. Умеренный выпас способствует сохранению структуры почвы, следовательно, и мезопедобионтов. Периодические затопления, палы, сенокос на пойменных лугах влияют на структуру педобионтов с проявлениями, обозначенными в позициях 4-7, что приводит к максимальному упрощению комплекса, но не влияет на его существование во времени.
Эффект от воздействия зависит, во-первых, от уязвимости конкретного компонента экологической ниши, от перехода его в разряд критичных. Все виды воздействия на педобионтов (за исключением, вероятно, контактных ядов и проникающей радиации) реализуются через изменения тех или иных условий обитания, что в свою очередь может приводить к лимитированию их топической и трофической ресурсной базы. Это, на фоне остального, неизбежно порождает конкурентное напряжение. Во-вторых, эффект определяется характером воздействия: интенсивностью (амплитудой), скоростью нарастания нагрузки и повторяемостью воздействия.
В связи с этим, в соответствии с общей концепцией Пригожина (2000), для сообществ мезопедобионтов в критических состояниях отмечаются три основных типа его динамики в зависимости от характера воздействия (рис.5). Тип А (сильное однократное воздействие, превышающее порог устойчивости биосистемы): необратимость процесса деградации первоначального сообщества (изменения плотности, видового состава, пространственного распределения, соотношения трофических групп, переориентация биотических связей, изменения функциональных характеристик, или биоценотической значимости). Тип Б (слабое краткосрочное воздействие): по причине достаточной силы связей между внутренними элементами сообщества, т. е. его упругости, происходят несущественные флуктуации значений тех же показателей вблизи точки устойчивости на время действия возмущающих факторов. Тип В (не превышающие порог устойчивости хронические воздействия): постоянное состояние нестабильности в связи с накоплением дозы приводит к нарастанию во времени амплитуды флуктуаций, разрыву внутренних связей между элементами сообщества и к неизбежному отклику в соответствии с законом «всё или ничего», в результате которого осуществляется переход сообщества в новое устойчивое состояние (теория Холинга) или его полное разрушение.
Сообщество педобионтов характеризуется низкой интегрированностью, по Шмальгаузену (1968). Подвижность компонентов, взаимозаменяемость, викарирование допускают более или менее автономные преобразования, что ведёт к росту устойчивости сообщества к внешней среде.
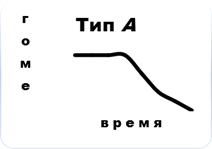
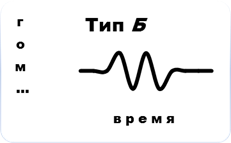
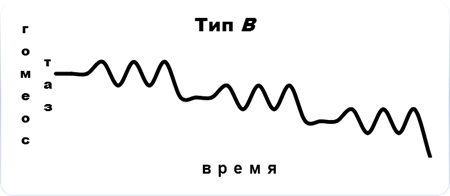
Рис. 5. Динамика сообщества мезопедобионтов в критических состояниях: тип А – сильное однократное воздействие, превышающее порог устойчивости биосистемы; тип Б – слабые краткосрочные воздействия; тип В – хронические воздействия, не превышающие порог устойчивости. По оси абсцисс и ординат – условные единицы.
Регуляция устойчивости, как показывают данные, осуществляется через структуру функциональных блоков сообщества. Снижение уровня устойчивости может проходить в несколько этапов: I (краткосрочные и обратимые изменения) – смена доминантов в гильдии/функциональной группировке, II (среднесрочные, часто обратимые изменения) – концентрирование доминирования в связи с ростом количества лимитирующих факторов, III (среднесрочные, редко обратимые) – монодоминирование в гильдии/группировке по той же причине при минимуме её устойчивости, IV (долгосрочные, чаще необратимые изменения) – полная элиминация видов, гильдий и группировок из функционального блока. Этапы отражают переориентацию биотических связей по причине уменьшения объёма доступных ресурсов и ужесточения условий их эксплуатации, приводящую к разрушению ценоза. При этом принципиальных различий между природно-климатическим и антропогенным происхождением нарушений нет.
Наиболее чувствительными мезопедобионтами к иссушению, изменениям кислотности гумусированного горизонта и механической структуры почвы являются диплоподы полизонииды Angarozonium amurense, геофиломорфные многоножки Escarius japonicus, фитосапротрофные брюхоногие моллюски Discus pauper, земляные кольчатые черви Eisenia nordenskioldi pallida и Drawida ghilarovi. Эти виды беспозвоночных по параметрам плотности могут быть биоиндикаторами антропогенных изменений таких параметров в среде обитания в условиях региона.
Все методы биологической диагностики почв основаны на понимании того, что почва как среда обитания составляет единую систему с населяющими ее популяциями разных организмов. В зависимости от сочетания природных факторов, определяющих почвообразовательный процесс, разные почвы различаются по составу своей биоты. Знание закономерностей взаимоотношений живых организмов с почвенной средой позволяет более корректно использовать биоту в качестве диагностов состояния природных и антропогенно-нарушенных экосистем или их отдельных компонентов.
Контроль экологического состояния окружающей среды на основе биоиндикаторов (индикационных признаков, отдельных видов или популяций) обладает целым рядом преимуществ:
- в условиях хронических антропогенных нагрузок биоиндикаторы могут реагировать на относительно слабые нагрузки вследствие эффекта кумуляции дозы;
- биоиндикаторы суммируют действие всех без исключения биологически важных факторов антропогенного воздействия и отражают их влияние на состояние окружающей среды в целом, фиксируют скорость происходящих в ней изменений, раскрывают тенденции развития экосистем, позволяют контролировать их состояние без необходимости постоянной регистрации химических и физических параметров, характеризующих качество среды, указывают места скопления в экосистемах различных загрязняющих веществ и токсикантов.
Выбор отдельных видов, популяций или сообществ, а также уровня организации биологических систем для диагностики состояния почвенной среды зависит от:
1. Типа стрессора (физический, химический, биологический).
2. Типа индикации (ранняя, прогнозная, сопутствующая, предваряющая).
3. Периода действия стрессора.
4. Способности почвенной системы противостоять нагрузкам.
Биоиндикационный исследования необходимо проводить в динамике, с использованием абсолютных и относительных стандартов сравнения и основой должен быть комплексный подход.
По степени роста чувствительности биоиндикаторы загрязнения почвы ранжируются в ряд: многоножки, моллюски, дождевые черви, паукообразные, мокрицы. Почвенные насекомые мало чувствительны к загрязнению почвы тяжелыми металлами и могут не рассматриваться в качестве биоиндикаторов загрязнения почвы. Выявлена сильная корреляционная связь между показателем жизненности беспозвоночных почвенных животных – биоиндикаторов загрязнения почвы – к индексу химического загрязнения почвы.
1. Бирюкова Н.А. Основы экологии.- М: Видос, 2004.
2. Муравьёв А.Г., Пугал Н.А., Лаврова В.Н. Экологический практикум, учебное пособие. С-Пб.: Крисмас+, 2007.
3. Экологические проблемы: что происходит, кто виноват и что делать?: Учебное пособие/Под ред. проф. В.И. Данилова-Данильяна. М.: Изд-во МНЭПУ, 2007.
4. Самедов П.А. – Изменение физических свойств почв деятельностью беспозвоночных животных. 17-й Международный конгресс почвоведов, том.1, 2008 г, Таиланд.
5. Самедов П.А., Надиров Ф.Т. – Влияние дождевых червей и мокриц на физико-химические и поверхностные свойтсва почв. Ж. Почвоведение, № 8, 2006, с.109 – 115.
6. Смирнов Ю.Б. Изучение почвенной мезофауны целинных степей с целью биоиндикации загрязнения почв тяжелыми металлами.
7. Степановских А. С., Охрана окружающей среды, Москва: «Юнити», - 2007
8. Шустов С.Б., Шустова Л.В. Химия и экология. – Нижегородский гуманитарный центр. 2004.
9. Экологическое образование: (Сборник программ по курсу экологии). – Нижний Новгород, 2008.
10. www.eco.priroda.ru
11. www.securpress.ru
12. www.docyment.ru
Перепечатка материалов без ссылки на наш сайт запрещена