Дипломная работа: Структурно-функціональні особливості наднирників людини на різних етапах онтогенезу
Дипломная работа: Структурно-функціональні особливості наднирників людини на різних етапах онтогенезу
Міністерство освіти та науки України
Прикарпатський національний університет імені Василя Стефаника
Інститут природничих наук
Кафедра біології та екології
Зав. кафедри д.б.н. проф. В.І.Парпан
Структурно-функціональні особливості наднирників людини на різних етапах онтогенезу
(дипломна робота на здобуття кваліфікації спеціаліст-біолог)
Виконала:
студентка 5 курсу групи Б-51
Савич Віталія
Науковий керівник:
д.б.н. проф. Грицуляк В.Б.
Івано-Франківськ-2009
Зміст
Вступ
Розділ І Огляд літературних джерел
Розділ ІІ Матеріал і методика дослідження
Розділ ІІІ Результати дослідження та їх обговорення
3.1 Загальні відомості про наднирникові залози
3.2 Гістогенез та ембріогенез наднирників
3.3 Патологічні зміни наднирників
Висновки
Список використаної літератури
Вступ
Актуальність теми. В системах і механізмах, з допомогою яких відбувається реалізація спадкової програми в ході індивідуального розвитку організму, важливе місце займає гормональна регуляція. В процесі онтогенезу в механізмі формування захисних реакцій організму велике значення має фізіологічна система гіпофіз – наднирники, в якій важливу роль відіграє кіркова речовина наднирників.
Історія вивчення наднирників налічує більше 300 років. За цей час накопичена обширна інформація про їх будову, функції і розвиток. Встановлена виключно важлива роль цих залоз у підтриманні сталості внутрішнього середовища організму і підвищення його опірності. Перш за все це зумовлено широким спектром біологічної дії кортикостероїдних гормонів на організм ссавців, ведучою роллю їх в процесах, що контролюють основні функції клітини. Біологічний ефект кортикостероїдних гормонів проявляється в суттєвій зміні в обміні речовин ссавців. Встановлення змін, що настають протягом життя в структурі наднирників, в характері їх взаємодії з гіпоталамусом та гіпофізом, а також з ефекторними органами і тканинами, вивчення специфіки зворотних зв’язків та зміни їх з віком дає цілісне уявлення не лише про вікові порушення в самій залозі, а і про роль їх в загальній системі нейрогуморальної регуляції обміну і функцій.
В процесі онтогенезу відбуваються глибокі зміни як в загальній масі, гістологічній структурі, так і фізіологічній активності кіркового шару. Змінюється співвідношення між кірковим і мозковим шарами наднирників ссавців.
Мета і задачі дослідження
Мета дослідження: виявити структурно-функціональні особливості наднирників людини на різних етапах онтогенезу.
Для виконання мати нами було поставлено наступні завдання:
1. Вивчити структурно-функціональні особливості наднирників у ембріонів та дітей;
2. Дослідити наднирники в процесі старіння у зрілих та похилого віку особин; патологічні стани наднирників;
3. Описати отримані результати.
Наукова новизна одержаних даних. Пізнання становлення регуляторних функцій і виявлення їх ролі в різноманітних процесах онтогенезу складають один з найактуальніших розділів біології розвитку. В останні роки збільшилася кількість наукових робіт, що присвячені питанням формування гормональних механізмів, а також вияснення їх функціональної ролі в процесі раннього онтогенезу організму, що стало можливим завдяки розробкам і застосуванню нових експериментальних методів дослідження. Останнім часом ведуться інтенсивні дослідження наднирників у зародків людини та ссавців. Дослідників цікавить не лише динаміка розвитку органа, але і становлення його функції. У виясненні функціональних можливостей надниркових залоз, їх ролі в забезпеченні адаптації плоду до мінливих умов існування зацікавлені як представники теоретичних дисциплін, так і практичні медики, що займаються проблемами охорони здоров’я. В роботі розглядається розвиток наднирників і встановлення їх функції у зародків ссавців та людини, процеси та особливості старіння їх.
Практичне значення одержаних результатів.
Одержані результати можуть слугувати для подальшого вивчення онтогенезу наднирників, використовуватися як навчальний матеріал для студентів.
Розділ І Огляд літературних джерел
ендокринологія гормон наднирник ембріон
Історія вивчення наднирників налічує більше 300 років. За цей час накопичена обширна інформація про їх будову, функції і розвиток.
Виникнення ендокринології як науки відноситься до середини ХІХ ст., коли в 1849 р. Бертольд вперше показав, що підшивання сімяників в черевну порожнину півням після їх кастрування попереджує у них розвиток посткастраційного синдрому. В цьому ж році С.Е. Броун-Секар, видаливши наднирники у тварин, доказав життєву необхідність цих залоз. Ендокринологія розвивалася під впливом передових ідей С.П. Боткіна, І.М. Сєченова, І.П. Павлова. В ній формувалося уявлення про цілісність організму в тісному взаємозв’язку нервової та ендокринної систем. Серед перших робіт, що присвячені вивченню внутрішньої секреції, була докторська дисертація Д.Галузинського "Про значення наднирникових тіл в тваринному організмі", що надрукована в 1862 р. Автор проводив адреналектомію у експериментальних тварин, вивчав взаємозв’язок наднирників з іншими залозами внутрішньої секреції.
Важливе значення для розвитку експериментальної ендокринології мали дослідження В.В.Підвисоцького. Він відмітив роль наднирників в регуляції обміну речовин, діяльності судинної та нервової систем (1896 р.). А.А. Богомолець у своїй докторській дисертації "До питання про мікроскопічну будову і фізіологічне значення наднирникових залоз в здоровому та хворому організмі" (1909 р.) вперше вказав на ліпоїдну природу гормонів кори наднирників. В 1936 р. канадський вчений Ганс Сальє створив вчення про загальний синдром адаптації. Одним з суттєвих недоліків концепції є абсолютизація ролі АКТГ і гормонів кори наднирників в реакції організму на стрес практично без урахування ролі нервової системи та інших систем організму в цілому.
Неоціненний вклад в розвиток ендокринології вніс В.Д.Шервинський. він перший висловив думку про важливу роль гормонів наднирників в пристосувальних реакціях організму. Він також значно розширив поняття про патогенез і клініку акромегалії і запропонував застосовувати гормони при цьому захворюванні.
Надзвичайно важливим є значення для розвитку світової ендокринології роботи Н.А.Юдаєва і його школи. Фундаментальні дослідження, проведені Юдаєвим, дозволили внести уточнення в схему біосинтезу кортикостероїдів під впливом різноманітних факторів на організм і за різних ендокринних порушень.
При розвитку наднирників у людини в ранньому ембріональному періоді зачаток кори (первинна кора) поділяється на дві частини: тонку, вузьку – дефінітивну і масивнішу – фетальну кору [4]. Довгий час вважали, що наявність фетальної кори характерна лише для приматів, але потім її виявили і в інших ссавців: у броненосця, кішки [22]. Фетальна кора була виявлена також у леопардів, тигрів, морських левів, овець, свиней, кроликів. Далі фетальна кора іволюціонує, що може відбуватися в різних видів у різні строки – в пре- чи постнатальному періоді [26].
Але у деяких видів тварин інволює не вся фетальна кора, частина її елементів залишається і включається в дефінітивну [26]. Крім того, дослідження фетальної кори у кроликів показали, що у цього виду фетальна кора взагалі взагалі ніякої інволюції не переживає, але цілком включається в дефінітивну [4].
Не дивлячись на велику кількість літературних даних, присвячених фетальній корі [23], функціональне значення її до цього часу залишається загадковим [4]. Вважається, що фетальна кора виконує особливу роль, необхідну для ембріонального періоду [10]. Деякі автори [4] розглядають зародкову кору як місце утворення прогестерону або андрогенів, які неначе охороняють плід від естрогенів [26], що збільшуються в період вагітності [11]. Багато авторів виявили, що зародкова зона, відділена від дефінітивної кори, здатна здійснювати ті ж процеси біосинтезу, що і весь наднирник [22].
Дослідження Т.С.Сахацької і В.І.Алтухової [3] встановили, що як зародкова, так і постійна кора здатна перетворювати ендогенні субстрати в дегидроепіандростерон сульфат і гідрокортизон. Але швидкості утворення цих речовин в постійній і зародковій корі різні.
У зародка 16,5 днів, за даними Йосимовича [15], наднирники вже інкапсульовані, паренхіма складається з еозинофільних епітеліоїдних клітин, ядра масивні, міхурчасті [26]. На цій стадії в клітинах кори виявляються невеликі суданофільні крапельки [15].
Наднирники плоду людини мають крупні розміри, їх відносна вага в 10 – 20 разів більша, ніж у дорослих [4]. Кора залози складається з двох різних частин – постійної і зародкової зон. Наднирники у ембріона людини виникають на рівні між 7-м шийним і 2-м грудним нервами [6].
Санто (1974) знайшов, що на стадії 7 тижнів ембріонального розвитку в глибоких частинах примордіальної кори є клітини з добре розвинутою гладкою ендоплазматичною сіткою [1]. В цей період мітохондрії ще не мають крист тубулярного типу, вони починають виявлятися тільки на 10-му тижні ембріонального розвитку [4]. До цього терміну автор відносить початок стероїдогенезу в наднирниках людини. Аналогічні зміни можна простежити в плодах кролика в період з 19-го по 24-й день розвитку і розглядаються як важливий показник завершення цитодиференціації фетальних адренокртикальних клітин в стероїдогенні [22].
Вікові зміни в структурі наднирників виражаються в розростанні сполучної тканини, в частому розвитку мікро аденом, вогнищевих гіперплазій, лімфоплазмоцитарних інфільтратів, атрофій кіркової речовини наднирників [14].
Деякі автори [4, 22] при вивченні ультраструктурних змін кори наднирників в процесі старіння виявили фестончатість контурів ядер ендотеліальних клітин капілярів наднирників, посвітління матрикса мітохондрій, появу в деяких ендотеліальних клітинах вторинних лізосом, ліпідних гранул, вакуолізацію і дискомплексацію крист мітохондрій, гідратацію цитоплазми і внутріклітинних органоїдів ендотелію, що свідчило про дистрофічні процеси в капілярах наднирників старих тварин, порушення умов транскапілярного обміну [16, 17].
Кортикостерон був першим кортикостероїдом, який вдалося виділити з екстракту кори наднирників (1933). Потім був отриманий ще ряд стероїдних сполук. Всі вони є похідними циклопентанопергідрофенантену, до якого приєднані радикали, залежно від числа атомів вуглецю в них гормони кори наднирника поділяються на три групи сполук: сполуками з 21 атомів вуглецю, з 19 атомами і з 18 атомами. Останні дві групи – це статеві гормони (відповідно андрогени і естрогени), а з 21 атомами – власне кортикостероїди, що мають специфічну для гормонів кори наднирників біологічну дію.
З великої кількості сполук лише три кортикостероїди мають найбільшу фізіологічну активність і постійно визначаються в крові, що відходить від наднирника: кортизон, кортикостерон і альдостерон. Решта сполук, що мають певну біологічну активність, є попередниками цих гормонів чи метаболітами.
Найбільше продукується кортизолу, потім кортикостерон і лише біля 200 мкг альдостерону. Альдостерон відносять до мінералокортикоїдів (він приймає участь в регуляції вуглеводного, білкового і жирового обміну). При цьому кортикостерон займає в певній степені проміжне положення, оскільки являється глюкокортикоїдом, і виявляє досить виражені мінералокортикоїдні властивості.
Альдостерон продукується в клубочковій зоні кори наднирників, а кортизол – в пучковій. В сітчастій зоні продукується група наднирникових статевих гормонів – андрогенів і естрогенів. В нормі естрогени виділяються наднирниками в кров у мінімальній кількості. У великій кількості продукуються андрогени. Андростендіон має найбільшу андрогенну активність; 11-оксиаандростендіон – найменшу. В невеликій кількості в крові надниркової вени спостерігається прогестерон. Він є одним з проміжних продуктів в біосинтезі кортикостероїдів; його не відносять до істинних гормонів наднирників. Допускається можливість утворення тестостерону, хоча головне місце біосинтезу останнього – гонади.
Завдяки зусиллям багатьох дослідників біосинтез кортикостероїдів вивчений порівняно повно. Першою стадією кортикостероїдогенезу є перетворення вихідного продукту – холестерину в прегненолон, який потім перетворюється в прогестерон. Якщо прогестерон першочергово піддається гідроксилюванню з 21-го атома вуглецю, то кінцевим продуктом буде кортикостерон. З 17-оксипрогестерону і 17-оксипрегненолону утворюються андрогени. Попередником альдостерону є кортикостерон. В процесі утворення кортикостероїдів використовується велика кількість НАДФ * Н, який утворюється завдяки посиленому обміну глюкози в наднирниках в пентозному циклі.
Біосинтез гормонів кори наднирників забезпечується дією специфічних ферментів – гідроксилаз і дегідрогеназ.
Після потрапляння в кров велика частка кортикостероїдів зв’язується з білками плазми. У результаті такого комплексування зв’язаний гормон ніби виключається з сфери фізіологічної дії і метаболізму, а біологічна ефективність гормону визначається концентрацією його вільної фракції.
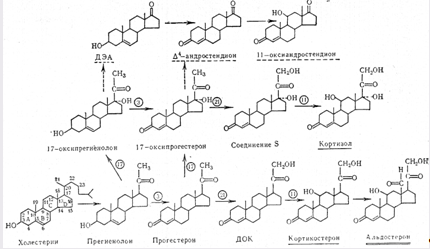
Схема біосинтезу гормонів кори наднирників
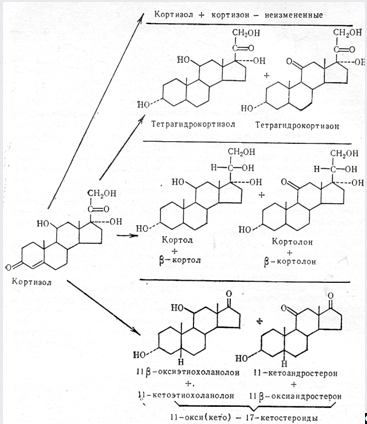
Головні метаболіти кортизолу
Виділення кортизолу протягом доби відбувається нерівномірно. Найбільше його виділяється о 6 годині, а найменше – з 20 до 24 год. Добовий ритм виділення кортизолу зумовлений добовим ритмом коливань рівня АКТГ в крові. Виробництво кортизолу повністю знаходиться під контролем АКТГ. Механізм дії АКТГ – це активація перетворення АТФ в циклічний 3,5-аденозинмонофосфат.
Секрецію АКТГ стимулює кортикотропін-рилізинг-фактор. Секреція альдостерону підвищується під впливом юкстагломерулярного апарату нирки, гіперкаліємії, гіпонатріємії.
Секреція адреналіну та норадреналін регулюється симпатичною нервовою системою і вищими центрами, що розташовані в корі головного мозку, ретикулярній формації та гіпоталамусі. Утворення адреналіну посилюється під впливом глюкокортикоїдів. Після потрапляння у кров основна кількість адреналіну та норадреналін зв’язується з альбуміном, а менша частина залишається у вільному стані.
Адреналін посилює серцеві скорочення, прискорює пульс, підвищує артеріальний тиск. Адреналін також розслаблює гладку мускулатуру бронхів, кишечника. Він розширює судини м’язів і серця та звужує судини шкіри, слизових оболонок і органів черевної порожнини. Адреналін сприяє скороченню м’язів матки і селезінки. Він бере участь в пігментному обміні, підвищує чутливість щитовидної залози до дії ТТГ.
На відміну від адреналіну норадреналін майже не впливає на вуглеводний обмін і гладку мускулатуру. Він підвищує артеріальний тиск в основному за рахунок діастолічного, що зумовлено його здатністю звужувати артеріоли. Поряд з адреналіном та норадреналіном має біологічну активність і дофамін. Адреналін, норадреналін та дофамін називають катехоламінами.
Розділ ІІ Матеріали і методи дослідження
Робота виконана на 12 препаратах наднирників людей різного віку забраних у паталогічному відділенні обласної клінічної лікарні.
Основним етапом вивчення зразків було дослідження їх у класичному світловому мікроскопі. Матеріал фіксували з наступним виготовленням забарвлених зрізів.
Досліджений об’єкт з самого початку фіксують. Фіксатори денатурують білки, стабілізують структури. Нами використовувалися такі фіксатори як формалін, спирт, оцтова кислота.
Після фіксації і промивання у воді препарати наднирників занурювали послідовно у спирти зростаючої концентрації (від 50 до 100 %) для обезводнення та ущільнення матеріалу, а далі у ксилол або бензол для просвітлювання.
На наступному етапі препарати наднирників просочували пластичною речовиною (парафіном). Після такого просочування їх наклеювали на дерев’яні колодочки і за допомогою мікротому робили гістологічні зрізи. Товщина їх може коливатися від 2 до 10 мкм.
Готові зрізи наклеювали на предметне скло за допомогою яєчного білка з гліцерином. Далі з них видаляли парафін, проводячи через ксилол або толуол. Потім препарати пропускали послідовно через етилові спирти, концентрація яких зменшується, промивали у воді і фарбували. Пофарбовані зрізи знову промивали у воді, пропускали через етиловий спирт і заключали у канадський бальзам – смола хвойного дерева, яка добре розчиняється у ксилолі.
Розділ ІІІ Результати дослідження та їх обговорення
3.1 Загальні відомості про наднирникові залози
Права надниркова залоза має трикутну форму, ліва – півмісяцеву. Надниркова залоза – орган парний, маса одної залози людини 12 – 13 г. Нижній край наднирників дещо роздвоєний і охоплює нижні полюси нирок ("шапка Наполеона"), розміщуючись разом з нирками в спільній жировій клітковині. Вони знаходяться на рівні від ХІ грудного до І поперекового хребця. Правий наднирник своїм верхнім краєм прилягає до печінки, а лівий вільний і з передньої поверхні покритий парієнтальною очеревиною. Розміри наднирників людини можуть сильно варіювати, але в середньому їх довжина становить 45 мм, ширина – 25 – 30 мм, товщина 6 – 10 мм.
Розрізняють три поверхні надниркової залози: передню, задню і нижню. На передній поверхні розміщені ворота (hilus). Зовні надниркова залоза вкрита фіброзною капсулою. До фіброзної капсули прилягає кора, яка складається з трьох зон: клубочкової, пучкової та сітчастої. В центрі надниркової залози знаходиться мозок, утворених великими клітинами двох видів: епінефроцити (основна маса мозку), норепінефроцити (розсіяні в мозку невеликими групами). У людини і ссавців мозок завжди оточений кірковою паренхімою. Але у інших тварин розташування його може бути іншим. Наприклад, у рептилій та птахів ці шари перемішані. У земноводних вони знаходяться поруч, а в риб і амфібій розташовані зовсім окремо. Досліджуючи розвиток наднирників у філогенезі, неважко помітити, що є тенденція до зближення інтерреналової і хромафінної тканини і проникнення останньої всередину інтерреналової. В організмі людини також можна знайти інтерреналову тканину, необ’єднану в комплекс з хромафінною. Це так звані додаткові тільця, які частіше складаються з кори або мозку, і лише в 16 % випадків – з обох структур, нагадуючи наднирники в мініатюрі. Додаткові інтерреналові тільця виявлені в сполучній тканині, що оточує наднирники, у верхніх відділах нирок, сонячному сплетінні та інших відділах черевної порожнини. Хромафінова тканина у людини поширена ще більш широко і може бути дифузно розселена по всьому організмі.
Кора розвивається з мезодерми ( з інтерреналової тканини, розміщеної між двома первинними нирками). Мозок розвивається з ектодерми, з ембріональних нервових клітин – симпатобластів.
Кора виробляє гормони – кортикостероїди, які поділяються на три групи:
- мінералокортикоїди (альдостерон) – виробляються клітинами клубочкової зони;
- глюкокортикоїди (гідрокортизон, кортикостерон, 11-дегідро- і 11-дезоксикортикостерон – виробляються в пучковій зоні;
- статеві гормони (андрогени, естроген, прогестерон) – виробляються клітинами сітчастої зони.
Наднирник ззовні покритий сполучнотканинною капсулою, що відділяє його від навколишньої жирової клітковини. В області воріт з капсулою зростається адвентиція центральної вени. У внутрішньому шарі капсули поряд з колагеновими і еластичними волокнами залягають гладкі м’язові клітини, завдяки наявності яких капсула володіє відомою скоротливістю.
Кора наднирників побудована з епітеліальних тяжів, чи стовбурів, що орієнтовані перпендикулярно до капсули. Стовбури тісно прилягають один до одного і розділені тонкими прошарками сполучної тканини, які відходячи від капсули, складають своєрідний каркас, або строму наднирника. По цих сполучнотканинних прошарках проходять кровоносні судини, що обплітають стовбури, і нервові волокна.
Кожен стовбур складається з декількох рядів епітеліальних клітин, форма, величина і положення яких відрізняється на різних рівнях стовбура. У верхніх кінцях стовбурів залозисті клітини, невеликі за розмірами і злегка приплюснуті, або нашаровуються один на одного, або їх ряди дугоподібно згинаються. Така будова верхніх ділянок стовпів дозволяє назвати цю частину кори клубочковою зоною. В наднирнику людини кубочкова зона представлена неправильними скупченнями дрібних епітеліальних клітин. Безпосередньо під капсулою залягає шар дрібних недиференційованих епітеліальних клітин, в яких часто трапляються мітози, особливо під час регенерації кори наднирників після її часткового порушення. Одночасно мітози спостерігаються також у верхніх клітинах клубочкової зони.
Якщо капсула видалена, то регенерація кори не відбувається. Навпаки, якщо ізольовану капсулу разом з невеликою кількістю епітелію клубочкової зони трансплантувати в інше місце, то настає проліферація кортикального епітелію. Джерелом цієї проліферації є вузький прошарок недиференційованих епітеліальних клітин, що залягає безпосередньо під капсулою. За рахунок проліферації клітин даної субкапілярної бластеми можуть виникати додаткові інтерренальні тільця, що нерідко спостерігаються в навколонаднирниковій клітковині.
Клітини клубочкової зони, багатокутної чи дещо заокругленої, неправильної кубічної форми, відрізняються злегка ущільненою цитоплазмою і невеликими овальними мітохондріями. В цих клітинах містяться дрібні вакуолі і порівняно небагато невеликих ліпоїдних крапель.
Мозок виробляє гормони – адреналін і норадреналін. Адреналін зменшує запаси глікогену в м’язах, збільшує вміст вуглеводів у крові, посилює і прискорює скорочення серцевого м’яза, звужує судини, підвищує артеріальний тиск. Адреналін та норадреналін за своєю хімічною природою здатні енергійно відновлювати оксиди важких металів. Тому при обробці наднирника розчином двохромовокислого калію медулярна частина зафарбовується бурим забарвленням внаслідок випадання нижчих оксидів хрому. У зв’язку з цим клітини мозкової частини отримали назву хром афінних чи феохромних. Ці клітини порівняно великі за розмірами, округлі, багатокутні чи призматичні. В їх цитоплазмі знаходяться багато чисельні гранули, що огорнуті ліпоїдною оболонкою і заповнені дрібною зернистістю. Ці гранули містять адреналін та норадреналін. Але після виведення катехоламінів ці гранули зберігаються в цитоплазмі. Відповідно, дані включення є не гранулами секрету, а специфічними органоїдами хромафінної клітини, на яких відбувається біосинтез катехлорамінових секреторних продуктів.
Серед хромафінних клітин є дві різновидності: одні виробляють адреналін, інші – норадреналін. Залозисті клітини медулярної частини розміщуються невеликими тяжами чи округлими скупченнями і обернені своїми апікальними кінцями до апікальних синусоїдів.
Наднирники, що розташовані поряд з черевною аортою, рясно постачаються кров’ю. Надниркова залоза кровопостачається верхньою, середньою та нижньою наднирковими артеріями. Ввійшовши в наднирник, ці артерії розгалужуються в капсулі або в кірковій речовині, даючи сітку капілярів, що обплітають епітеліальні тяжі кіркової речовини. Стінки капілярів утворені поряд з ендотеліальними також ретикулярними клітинами, що мають здатність поглинати і накопичувати чужорідні колоїдні речовини. Мозкова частина наднирників отримує кров через особливі артерії, які пробурюють кору без галуження і розпадаються на капіляри між скупченнями і тяжами медулярних клітин. На межі кіркової і мозкової частини (в сітчастій зоні кори) капіляри розширюються і переходять в широкі венозні синусоїди мозкової частини. Ці синусоїди, стискаючись, дають початок венам, що збираються в центральну вену, яка виходить з наднирника і впадає в нижню порожнисту вену справа і в ниркову вену зліва. Венозна кров відтікає в однойменні вени. Частково кров з мозкової частини наднирника виноситься по особливих венах, які оминаючи кору, за межами наднирника виливають кров у вени ворітної системи. Таким шляхом адреналін і норадреналін переноситься у печінку (де впливає на мобілізацію глюкози). Капіляри верхніх шарів кори і капсули дають початок самостійній сітці вен, що впадають також в судини ворітної вени.
Лімфа відтікає в поперекові лімфатичні вузли. Лімфатична сітка наднирників потужна. Лімфатичні капіляри залягають в прошарках сполучної тканини між стовбурами кори і між клітинними тяжами в мозковій частині. Лімфатичні судини кори збираються в субкапсулярне сплетіння, що дає початок виносним стовбурам. В мозковій частині лімфатичні капіляри збираються в сітку, що оточує центральну вену, разом з якою виходять виносні лімфатичні судини.
Іннервація наднирників багата. Симпатична іннервація здійснюється черевним сплетінням, утвореним аксонами вісцеромотонейронів грудних і поперекових вузлів, в яких перериваються гілки блукаючого нерва. Основним джерелом іннервації є черевні нерви, подразнення яких посилює продукцію адреналіну в мозковій частині і його виділення у венозну кров. Навпаки, перетин черевних нервів приводить до послаблення діяльності мозкової частини.
Оскільки медулярна частина наднирників являє собою видозмінений симпатичний ганглій, волокна черевного нерва слід розглядати як прегангліонарні. Роль же постгангліонарних нейронів у цьому еферентному ланцюгу відіграють хромафінні клітини мозкової речовини. Їх гормони, досягаючи з кровообігом крові ефектора, впливають на нього збуджуючи чи гальмуючи, відповідно доносять до нього гуморальним шляхом імпульс. Що посланий спинним мозком.
Поряд з черевними нервами в іннервації наднирників беруть участь блукаючі і діафрагмальні нерви. Нервові волокна, що входять в наднирник, утворюють рясні сплетіння в капсулі, кірковій і медулярній частині. Від капсулярного сплетіння нервові волокна опускаються вздовж стовбурів кори. Найрясніше галуження нервових волокон в клубочковій і сітчастій зонах кори. Густе нервове сплетіння залягає також в медулярній частині. Дуже рясно інервовані судини, особливо центральна вена. По всьому наднирнику як в капсулярному сплетінні, так і в корі і особливо в мозковій частині розкидані скупчення симпатичних нервових волокон – мікро ганглії, а по ходу нервових волокон часто трапляються поодинокі нервові клітини. В корі наднирників можна побачити закінчення еферентних волокон на залозистих клітинах. А в мозковій частині волокна, що виходять з сплетіння, закінчуються на поверхні хроматофорних клітин невеликими кільцями чи петлями. В той же час наднирник відрізняється різноманітністю чутливих (рецепторних) нервових закінчень. Рецепторні структури представлені як простими ґудзиковими чи колоподібними закінченнями, так і складними деревоподібними рецепторами, що утворюють рецепторні поля і клубки, що залягають в кірковій частині навколо і між її стовбурами. В мозковій частині виявляються потужні рецепторні поля. Такий великий розвиток рецепторних утворень заставляє вважати, що аферентній сигналізації, що поступає з наднирників, повинна належати суттєва фізіологічна роль.
Нижче від клубочкової зони наявна вузька смужка, що складається з 3 – 4 рядів дрібних клітин з гомогенною базофільною цитоплазмою. Ліпоїдні включення в клітинах відсутні, тому ця смужка отримала назву суданофобної чи проміжної зони. Знизу середню частину стовбурів займає пучкова зона. Клітини цієї зони великі, кубічної чи багатокутної форми, але найбільше серед них призмоподібних. В клітинах клубочкової зони особливо сильно накопичуються ліпоїди і нейтральні жири, чим зумовлюється характерне жовте забарвлення, що властиве корі наднирників в нормі. Жиро-ліпоїдні включення мають вигляд округлих крапель різної величини.
Поряд з світлими спонгіоцитами в пучковій зоні, а також в наступній сітчастій зоні трапляються клітини з щільнішою цитоплазмою. Мітохондрії в клітинах пучкової зони значно багато чисельніші і крупніші, ніж в клітинах клубочкової зони. Внутрішня оболонка цих мітохондрій замість правильних крист утворює звивисті і галузисті диверкули, що сильно збільшують внутрішню поверхню. Ендоплазматична сітка в клітинах пучкової зони виражена слабко, але у цитоплазмі завжди трапляються вакуолі, які в клітинах нижньої частини пучкової зони і в сітчастій зоні мають вигляд крупних лакун чи так званих сумчастих утворів. Між клітинами і капілярами залишаються вільні щілини, що містяться також між сусідніми залозистими клітинами. У ці щілини входять мікроволоконця, що виступають над краєм залозистих клітин. Сюди ж відкриваються "сумчасті утворення", чим забезпечується швидке виділення секреторних продуктів.
Світлі і темні клітини представляють послідовні стадії секреторного циклу. Утворення секрету в корі наднирників відбувається всередині мітохондрій, і при розриві останніх гранули секрету потрапляють в цитоплазму. Світлі клітини знаходяться в стадії активної віддачі секрету. В результаті розпаду мітохондрій і виливання вмісту "сумчастих лакун" світлі клітини приймають вигляд темних. В темних клітинах мітохондрії утворюються заново, тому вони малі за розмірами і компактні. По мірі поступової гіпертрофії мітохондрій і скупчення "сумчастих лакун" темна клітина приймає вигляд світлої. Таким чином, в темних клітинах, очевидно, переважають процеси біосинтезу секрету.
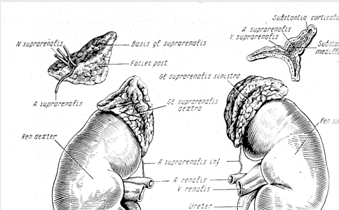
Мал. 1. Наднирники (за Ф.Кишш і Я. Сентогораї)
Інтенсивність адренокортикального гормоногенезу не виявляє зв’язку з кількістю ліпоїдних крапель, що накопичуються в цитоплазмі залозистих клітин пучкової зони. В нормі секреція кори наднирників здійснюється за мерокриновим типом, але під час посиленої секреції, що виникає за різкої потреби організму в адренокортикальних гормонах, в залозах клітин кори секреція інколи має голокриновий характер. В нижній частині кори пучкова зона переходить в сітчасту, що межує з медулярною частиною. В сітчастій зоні правильне стовбурове розташування залозистих клітин втрачається і епітеліальні тяжі розгалужуються, утворюючи пористу сітку.
Залозисті клітини, що досягають крупних розмірів в пучковій зоні, знову зменшуються в об’ємі, а їх цитоплазма дещо ущільнюється, тому на препаратах вони мають темне забарвлення. Серед них трапляються дегенеруючи клітини. Часто в клітинах сітчастої зони відкладаються зерна буро-жовтого пігменту.
Не дивлячись на чітку різницю в розмірах і формі клітин різних зон, співвідношення між зонами може змінюватися у відповідності з зсувами в функціональній активності кори наднирників і за різних патологічних станів. Зокрема, посилене виділення глюкокортикоїдних гормонів супроводжується значним набряканням клітин пучкової зони, тому її ширина в даних обставинах значно збільшується. Послаблення секреції глюкокортикоїдних гормонів проявляється редукцією пучкової зони, від якої зберігається тільки її середня частина, а нижні ділянки приймають вигляд сітчастої зони, яка в цих умовах розширюється.
В клубочковій зоні відбувається утворення мінералокортикоїдів, в пучковій – глюкокортикоїдів, а сітчастій зоні приписується головна роль в утворенні кортикоостеронів, що виявляють адреногенну дію. До того ж механізми, що контролюють гормоноутворення в кожній із зон, різноманітні. Але ж при всій своїй відносній самостійності окремих зон, вони зберігають відому структурну єдність та функціональну взаємодію, тому кора наднирників є цілісною системою.
Дві частини наднирників продукують гормони, що різко відрізняються за хімічним складом і фізіологічною дією в організмі, регуляція їх відбувається окремо.
Кіркова і медулярна тканини інколи можуть траплятися в організмі ізольовано, у вигляді так званих додаткових наднирників. Вони зазвичай складаються з адренокортикального епітелію і представляють собою ізольовані острівки, що подібні на клітини пучкової зони. Такі тільця трапляються в нирці, вздовж сечоводів, в печінці, придатках сім’яників, яєчниках, широкій зв’язці матки.
Очевидно, кіркова і медулярна частини знаходяться в певній взаємодії. За нормального стану організму ці частини функціонують більш-менш окремо, але за різких порушень стану організму, що викликаються дією сильних і надзвичайно сильних подразників, коли збереження і відновлення його рівноваги забезпечується мобілізацією всіх його ресурсів, настає одночасна і сучасна активація обох частин наднирника.
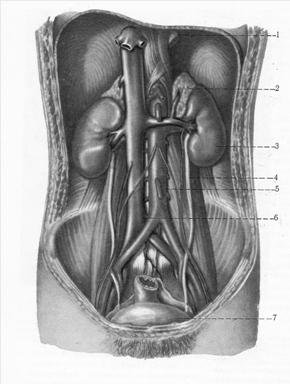
Фото 1. Топографія наднирників.
1-діафрагма; 2-лівий наднирник; 3-ліва нирка; 4-сечовід; 5-аорта; 6-вена; 7-сечовий міхур.
3.2 Гістогенез та ембріогенез наднирників
Наднирники як за походженням, так і за структурою, є складними залозами. Два компоненти залози - кора і мозок – є анатомічно і ембріологічно різними структурами. Джерелом розвитку кори наднирників є мезодермальний целомічний епітелій в районі пронефроса, тісно пов’язаний з закладками гонад. Джерелом тканини мозкової частини наднирника вважається ектодерма невральних валиків, з яких закладається вся вегетативна нервова система.
Закладка надниркових залоз відбувається дуже рано, у різних видів в різні терміни. При цьому частина клітин целомічного епітелію бічних пластинок розростається і входить в прилеглу тканину. В утворений інтерреналовий зачаток дуже скоро починають вростати тяжі клітин – хромафінобластів із зачатків симпатичних гангліїв. З моменту проникнення хромаффібринобластів в центральну частину інтерреналового зачатка поступово утварюється мозкова частина залози, або медула.
При розвитку наднирників у людини в ранньому ембріональному періоді зачаток кори (первинна кора) поділяється на дві частини: тонку, вузьку – дефінітивну і масивнішу – фетальну кору. Довгий час вважали, що наявність фетальної кори характерна лише для приматів, але потім її виявили і в інших ссавців: у броненосця, кішки. Фетальна кора була виявлена також у леопардів, тигрів, морських левів, овець, свиней, кроликів. Далі фетальна кора іволюціонує, що може відбуватися в різних видів у різні строки – в пре- чи постнатальному періоді.
Але у деяких видів тварин інволює не вся фетальна кора, частина її елементів залишається і включається в дефінітивну. Крім того, дослідження фетальної кори у кроликів показали, що у цього виду фетальна кора взагалі взагалі ніякої інволюції не переживає, але цілком включається в дефінітивну.
Не дивлячись на велику кількість літературних даних, присвячених фетальній корі, функціональне значення її до цього часу залишається загадковим. Вважається, що фетальна кора виконує особливу роль, необхідну для ембріонального періоду. Деякі автори розглядають зародкову кору як місце утворення прогестерону або андрогенів, які неначе охороняють плід від естрогенів, що збільшуються в період вагітності. Багато авторів виявили, що зародкова зона, відділена від дефінітивної кори, здатна здійснювати ті ж процеси біосинтезу, що і весь наднирник. Дослідження Т.С.Сахацької і В.І.Алтухової встановили, що як зародкова, так і постійна кора здатна перетворювати ендогенні субстрати в дегидроепіандростерон сульфат і гідрокортизон. Але швидкості утворення цих речовин в постійній і зародковій корі різні. Крім фетальної кори, яка спостерігається не у всіх ссавців, в адреналовій залозі зародкового періоду знайдені й інші додаткові зони. Кінцеву, дифінітивну будову наднирники набувають або на пізних стадіях пренатального періоду, або на початку постнатального, що залежить від виду тварин. А у деяких тварин (миші) це відбувається після статевого дозрівання. В дефінітивному стані кора наднирника складається з клубочкової, пучкової і сітчастої зон. Всі три зони мають чіткі межі і характеризуються певними функціями. Диференціація клубочкової зони в наднирниках зародків щурів спостерігається рано: вже на 16 – 17-й день розвитку чітко видно дрібні клітини, зібрані в клубочки, що лежать безпосередньо під капсулою наднирника. Але решта частини кори до народження не має чітко відмежованих границь між майбутньою пучковою та сітчастою зонами. У інших тварин, наприклад у кішки, зональна структура кори наднирника відмічена на 10 – 13-й день зародкового розвитку.
Поява зональності в корі наднирника іноді зв"язують з початком функціональної активності залози. Відомо, що навність ензимів стероїдного біосинтезу в клітинах наднирників щурів виявляється вже в 12-денних зародків, а в інкубаційному середовищі залози 13,5-денних зародків спостерігаються кортикоїди. В наднирнаках 8-денних плодів людини є необхідні для біосинтезу кортикоїдів ферменти. Здатність кори наднирників секретувати стероїди далеко опереджує морфологічну диференціацію залози.
Перші ознаки диференціації наднирників плодів щура спостерігаються на 11,5 днів розвитку, коли під поверхнею урогенітального валика з"являється локальне накопичення клітин. Стадія 13,5 днів зародкового життя характеризуються наявністю груп клітин з округлими, набряклими, багатоядерцевими ядрами і рясною гранулярною цитоплазмою, а також тісним зв"язком між клітинами, малою кількістю адреномодулярних клітин, відсутністю артерій і погано розвинутою капсулою. На стадії 15,5 днів навколо адреналової залози спостерігається конденсація мезенхіми, але чіткої капсули в цей час ще нема. Однак залоза вже легко відпрепаровується від навколишніх тктанин. У зародка 16,5 днів, за даними Йосимовича, наднирники вже інкапсульовані, паренхіма складається з еозинофільних епітеліоїдних клітин, ядра масивні, міхурчасті. На цій стадії в клітинах кори виявляються невеликі суданофільні крапельки. Останні у 17,5 денних зародків у внутрішній зоні, в області формування пучкової і сітчастої зон помітно збільшуються. За даними Алтухової В., в цей же час розвитку кора наднирника складається з пухких поліморфних клітин, в яких виявляються у невеликій кількості ліпіди, холестерин і аскорбінова кислота.
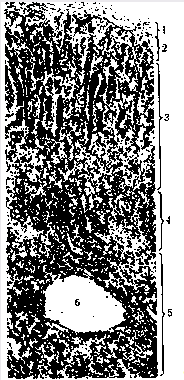
Фото 2. Зріз наднирника людини. 1-капсула; 2-клубочкова зона; 3-пучкова зона кіркової речовини; 4-сітчаста зона кіркової речовини; 5-мозкова речовина; 6-ворота наднирника.
У золотистого хом"ячка початок кортикостероїдної активності наднирника починається між 12,5- і 14,5-м днями розвитку. Залоза в цей період характеризується наявністю гладкого ендоплазматичного ретикулюму, мітохондріями з тубулярно-везикулярними кристами і активністю ферменту трибетагидроксистероїд дегідрогенази (фото 2).
Якщо поділити розвиток наднирника щура в пренатальному періоді на основні стадії, то їх буде чотири. Перша стадія характеризується незалежністю від ще не розвинутого гіпофіза, відбувається з 12,5 до 15,5 днів. В цей період наднирник продукує кортикостероїди, які спостерігаються в інкубаційному середовищі до 16-го дня розвитку. Друга стадія триває з 15,5 до 17,5 днів. В цей період кортикостероїди в інкубаційному середовищі не визначаються. Можливо, холестерин, що необхідний для синтезу кортикостероїдів витрачається на побудову мембран. Але якщо в інкубаційне середовище наднирника 17-денного зародка додати суспензію гіпофіза, то наднирник, хоч і слабо диференційований, набуває здатності до стероїдогенезу. Третя стадія, що складає період з 17,5 днів до народження, є стадією реактивності. В цей час спостерігається помітне збільшення розмірів наднирників, швидкість росту їх перевищує загальний ріст плода. Характерно, що збільшення об’єму наднирників відбувається за рахунок кори, тоді як медула в цей період складає лише 2 % від об’єму кортекса. АКТГ в гіпофізі щурів спостерігається з 18-го дня пренатального життя і в цей термін кортикостероїди виділяються як в самій залозі, так і в плазмі. Найбільший вміст їх визначається в 18-19-й день розвитку, тоді як до кінця вагітності кількість їх різко знижується. Четверта стадія, що настає після народження, характеризується відносною рефратерністю, зниженням секреції глюкокортикоїдів і зменшенням об’єму наднирників. Вага їх 4 години після народження падає з 2,56 до 2,27 мг, хоча вага тіла в цей час практично не змінюється.
Наднирники плоду людини мають крупні розміри, їх відносна вага в 10 – 20 разів більша, ніж у дорослих. Кора залози складається з двох різних частин – постійної і зародкової зон. Наднирники у ембріона людини виникають на рівні між 7-м шийним і 2-м грудним нервами. На стадії 8 мм розвитку (біля 21 доби) з’являються перші кортикальні клітини, які утворюють зародкову, або „фетальну", зону, а пізніше утворюється „дефінітивна" зона. На стадії біля 20 мм всередину кортекса починають проникати нервові елементи, що утворюють медулу, а на стадії біля 10 см відбувається завершення її формування, і до моменту народження вона займає незначний об’єм. Фетальна зона до моменту народження плоду досягає вершини свого розвитку і починає інволювати. Дефінітивна зона складається з декількох шарів однорідних клітин і розташовується безпосередньо під капсулою. Через декілька місяців після народження вона диференціюється в клубочкову, пучкову і сітчасту зони. За даними декількох авторів, зародкова зона, що складає до моменту народження 80 % об’єму залози, починає піддаватися процесу інволюції ще до народження, з 8-го місяця вагітності, а після народження цей процес посилюється.
В перші два тижні після народження малюка наднирники втрачають
50 % об’єму зародкової кори, а до кінця першого року життя вона складає біля 16 % залози. Ультраструктура клітин зародкової кори наднирників плоду в першому триместрі вагітності характеризується наявністю в цитоплазмі добре розвинутої ендоплазматичної сітки, щільно укладеними цистернами і вільними рибосомами. В постійній корі подібна структура з’являється лише у другій половині внутріутробного розвитку. Тонка структура зародкової кори свідчить про те, що ця зона вже з 8 тижнів зародкового розвитку володіє стероїдогенною активністю, а постійна зона цю властивість отримує лише з 6-го місяця пренатального розвитку.
Санто (1974) знайшов, що на стадії 7 тижнів ембріонального розвитку в глибоких частинах примордіальної кори є клітини з добре розвинутою гладкою ендоплазматичною сіткою. В цей період мітохондрії ще не мають крист тубулярного типу, вони починають виявлятися тільки на 10-му тижні ембріонального розвитку. До цього терміну автор відносить початок стероїдогенезу в наднирниках людини. Аналогічні зміни можна простежити в плодах кролика в період з 19-го по 24-й день розвитку і розглядаються як важливий показник завершення цитодиференціації фетальних адренокртикальних клітин в стероїдогенні.
Наднирники в процесі старіння
В процесі онтогенезу в механізмі формування захисних реакцій організму велике значення має фізіологічна система гіпофіз – наднирники, в якій важливу роль відіграє кіркова речовина наднирників. Перш за все це зумовлено широким спектром біологічної дії кортикостероїдних гормонів на організм, ведучою роллю їх в процесах контролюючих основні функції клітини. Біологічний ефект кортикостероїдних гормонів, введених ззовні або ендогенного походження, проявляється в суттєвих змінах в обміні вуглеводів, білків та жирів, в збереженні водно-сольового гомеостазу, в регуляції складних механізмів біосинтезу білка. Все це відображається у функціонуванні органів і систем, у формуванні синдрому адаптації. Встановлення змін, що відбуваються з віком в структурі і функції наднирників, в характері їх взаємодії з гіпоталамусом і гіпофізом (центрами регуляції), а також з ефекторними органами і тканинами, вивчення специфіки зворотних зв’язків і зміни їх з віком дають цілісну уяву не лише про вікові порушення в самій залозі, але і ролі їх в загальній системі нейрогуморальної регуляції обміну і функцій старіючого організму.
В процесі онтогенезу відбуваються глибокі зміни як в загальній масі, гістологічній структурі, так і в фізіологічній активності кори наднирників. Змінюється співвідношення між корою і мозком: у дорослих 1 : 1, а у старих 1 : 2. У чоловіків і жінок в постембріогенезі наднирники досягають максимальної маси (у жінок – 13 г, у чоловіків 13,9 г). В старості маса наднирників зменшується. У чоловіків після 70 років вона дорівнює 12,6, у жінок - 11,72 г.
У чотиримісячних плодів маса наднирників більша за нирки або маси їх однакові. Більшу частину залози в цей період займає кора. До восьмого року життя кіркова речовина наднирників досягає структурної зрілості і отримує морфологічну стійкість. В такому стані наднирники залишаються до старечого віку. Якщо в період молодості і зрілості особливо інтенсивно розвиваються клубочкова і сітчаста зони, то в процесі старіння ці зони кори наднирників інтенсивніше інволюють, ніж пучкова.
Вікові зміни в структурі наднирників виражаються в розростанні сполучної тканини, в частому розвитку мікро аденом, вогнищевих гіперплазій, лімфоплазмоцитарних інфільтратів, атрофій кіркової речовини наднирників. В клітинах кори наднирників також помітні дегенеративні зміни: нечіткі границі клітин, ядерні пікнози, поліхромазія і зморщування плазми, зменшення кількості кіркових клітин, нерівномірне розташування жиру в пучковій зоні, атрофія мозкового шару і розширення кровоносних капілярів.
Деякі автори при вивченні ультраструктурних змін кори наднирників в процесі старіння виявили фестончатість контурів ядер ендотеліальних клітин капілярів наднирників, посвітління матрикса мітохондрій, появу в деяких ендотеліальних клітинах вторинних лізосом, ліпідних гранул, вакуолізацію і дискомплексацію крист мітохондрій, гідратацію цитоплазми і внутріклітинних органоїдів ендотелію, що свідчило про дистрофічні процеси в капілярах наднирників старих тварин, порушення умов транскапілярного обміну.
Морфологічні дослідження кори наднирників у старих тварин показали різний ступінь змін їх в залежності від виду тварини. У старих мишей вони виражені значно більше ніж у щурів і птахів. В зрілому віці визначається також більша ширина пучкової і сітчастої зон кори наднирників.
Клубочкова зона кори наднирників продукує мінералокортикоїди. Основними гормонами, що мають біологічний ефект мінералокортикоїдного характеру, є альдостерон і 11-дезоксикортикостерон (ДОК). Альдостерон – найактивніший мінералокортикоїдний гормон клубочкової зони наднирників, необхідний для підтримання балансу рідини в організмі. Біологічно активний лише вільний альдостерон. Зв’язок гормону з білками дуже слабкий.
Наукові дані про вікову зміну обміну альдостерону в організмах ссавців протирічні. Деякі автори зазначають, що з віком рівень альдостерону в плазмі крові знижується. У жінок зрілого віку екскреція альдостерону з сечею більша ніж у чоловіків. Помітне зниження альдостерону у чоловіків відбувається після 80 років.
В клімактеричному періоді значно зменшується функція статевих залоз. Серед ссавців клімактеричний період притаманний для людей та китів.
Клініко-експериментальні дослідження вмісту альдостерону в плазмі крові в різні фази циклічних змін в організмі зрілого віку достовірно показали наявність зв’язку функціонування наднирників та гонад і в пізні терміни онтогенезу.
Пучковою зоною кори наднирників синтезуються і секретуються в кров гормони глюклкортикостероїдного ряду. Дія їх напрямлена на регуляцію обміну білків, жирів і вуглеводів. У підтримці гомеостазу в організмі в будь-яких стресових ситуаціях глюкокортикоїдні гормони мають переважне значення, тому що приймають участь у формуванні пристосувальних механізмів. В будь-якій ситуації, коли організму необхідно значну втрату гормону, він легко звільняється і переходить у вільний стан, в якому проявляє найбільшу біологічну активність.
При старінні розвиваються не тільки кількісні, але і якісні зміни в реакціях тканин на дію гормонів кори наднирників. Крім того, якісно змінюється реакція тканини наднирника на вплив специфічного стимулятора кортикотропіна. З віком росте реактивність, але зменшується реакційна здатність наднирника. Резервні можливості залози з віком знижуються оскільки при тривалій стимуляції або напрузі виявляється неадекватна реакція.
Концентрація рецепторів з великою схожістю з глюкокортикоїдами з віком знижується на одиницю маси органа і концентрацію білка цитозоля в м’язах та мозку.
У щурів в печінці і передміхуровій залозі концентрація цих рецепторів збільшується до 13 місяців, потім в печінці вона не змінюється, в передміхуровій залозі – зменшується.
Таким чином, глюкокортикоїдна функція кори наднирників в процесі старіння змінюється не тільки якісно, а й кількісно.
Сітчаста зона кори наднирників продукує андростероїди, які за хімічною структурою і прояву біологічної дії схожі з статевими гормонами. З сечею виділяються метаболіти андрогенів – нейтральні 17-кесостероїди (17-КС). По мірі старіння організму концентрація 17-КС в плазмі периферичної крові знижується, що відбувається поступово. Крім того, з віком знижується виділення дегідроепіандростерону і етіохоланолу. Андрогенна функція кори наднирників в процесі старіння страждає набагато раніше і більш виражено, ніж глюкокортикоїдна. Зменшення продукції андрогенів приводить до різкого зменшення їх концентрації в крові вже у 40-60 років. В старечому віці рівень андрогенів у крові важко визначити із-за малої концентрації його в крові. Андрогенна функція кори наднирників після 70 років зводиться до нуля.
Таким чином, андрогенна функція кори наднирників в процесі старіння знижується, а глюкокортикоїдна функція змінюється мало і на пізніших етапах онтогенезу.
При старінні на фоні збереженості базального рівня глюкокортикоїдів і зниження андрогенної функції кори наднирників виникає чутливість залози до кортикотропіну, падає реакційна здатність, потенційні резерви, скоріше наростає виснаження її діяльності. Ріст чутливості залози в старості до коортикотропіну може розцінюватися як пристосувальна реакція організму. Зниження реакційної здатності до тропного гормону обмежує адаптаційні можливості системи.
При щоденному повторенні стресів старі щурі гинуть швидше ніж молоді за таких же умов. Про більш ранній розвиток у старих тварин фази виснаження свідчить також стрімкий розвиток у них зниження екскреції кето стероїдів.
Мозкова речовина наднирника є автономною його частиною у функціональному відношенні. Секрет мозкової речовини наднирників може відразу потрапляти в печінку і викликати біологічну дію. Адреналін синтезується мозковою речовиною.
Мозкова речовина наднирників впливає на процеси старіння організму. При дії адреналіну в багатьох випадках у старих тварин зростає чутливість і падає реакційна здатність тканин. При введенні малих доз гормону зміщення в рівні цукру в крові, артеріальному тиску, тонусі судин кінцівок і нирок, збудженні рецепторів судин, рості висот скорочення скелетної мускулатури, зміни гліколізу, глікогенолізу, окислювального фосфорилювання в серці більш значимі в старих тварин. При введенні великих доз адреналіну функціональні і обмінні зрушення в багатьох випадках більш виражені у молодих тварин. При старінні знижується синтез медіатора, росте чутливість адренорецепторів.
Таким чином, в процесі старіння відмічаються неоднозначні зміни у функції різних шарів і зон кори наднирників. При фізіологічному протіканні процесів старіння (навіть до пізніх термінів онтогенезу) наднирники є надійною ланкою в здійсненні важливих механізмів адаптації.
3.3 Патологічні зміни наднирників
Адісонова хвороба
Адісонова хвороба – хронічна недостатність кори наднирників, що розвивається внаслідок двостороннього ураження цих залоз або недостатньої стимуляції наднирників адренокортикотропного гормону на ґрунті гіпоталамо-гіпофізарних порушень. Залежно від ураження розрізняють первинну (периферичну) і вторинну (центральну) недостатність кори наднирників. Захворювання зустрічається дуже рідко.
Вперше цей патологічний стан був описаний в 1855 р Адісоном. Етіологічним фактором первинної хронічної недостатності кори наднирників в більшості випадків (75 – 80 %) є туберкульоз наднирників, дуже рідко – сифіліс. Стерті форми захворювання з слабко вираженою симптоматикою можуть бути зумовлені будь-яким фактором, що викликає фактор напруження: стрес, інфекційні захворювання, отруєння, опіки, фізичне і розумове перевантаження, гіповітаміноз, психотравма.
Хронічна недостатність кори наднирників може розвиватися в результаті безпосереднього ураження кори наднирників (первинний гіпокортицизм) або внаслідок ураження гіпофізарної області і зменшення виробітку кортикитропіну з наступною гіпофункцією, гіпоплазією і атрофією кори наднирників (вторинний гіпокортицизм).
Первинна хронічна недостатність кори наднирників розвивається в результаті фіброкавернозного туберкульозу наднирників. Значно рідше туберкульозний процес локалізується лише в корі наднирників. Рідше захворювання пов’язане з амілоїдною дегенерацією кори наднирників, трормбозом їх судин, гемораргіями, ураженням при гнійних процесах, сифілісом, метастазами злоякісних новоутворень.
Хронічна недостатність кори наднирників може виникнути при тривалій кортикостероїдній терапії при різноманітних захворюваннях (ревматизм, інфекційний поліартрит, бронхіальна астма, хвороби крові), що пов’язано з гіпоплазією і атрофією кори наднирників із-за функціональних і органічних змін аденогіпофіза.
В основі патогенезу захворювання є різке зниження продукції стероїдних гормонів, внаслідок поширених деструктивних змін в корі наднирників. Зниження або випадіння функції клубочкової зони кори наднирників, що продукує мінералокортикоїди, викликає характерне для цього захворювання порушення водно-сольового обміну (гіпонатріємія, гіперкаліємія, гіпернатріурія і гіпокаліурія), збіднення організму хлоридами, обезводнення організму, зниження артеріального тиску. Ураження пучкової зони, що виробляє глюкокортикоїди, приводить до збіднення глікогеном печінки та м’язів, гіпоглікемії, схуднення, адинамії і зниження опірності організму до зовнішніх несприятливих факторів.
В результаті деспруктивних процесів в корі наднирників порушується утворення мінералокортикоїдів, глюкокортикоїдів і андрогенів. Дефіцит гормонів викликає складні і різні порушення багатьох обмінних процесів, що в свою чергу веде до порушення функції багатьох органів і систем.
Недостатність мінералокортикоїдів супроводжується в основному посиленою втратою натрію і, як наслідок, її дегідратацією, зменшенням об’єму циркулюючої крові. Втрата натрію і зменшення його вмісту в стінках дрібних судин знижує пресорну дію вазоактивних речовин, в першу чергу норадреналіну, що веде до зменшення периферичного опору, зниженню артеріального тиску.
Недостатність глюкокортикоїдів обтяжує наявні водно-електролітичні зміни, поряд з цим веде до глибоких змін вуглеводного, білкового та інших видів обміну. Послаблюються процеси неогліколізу, розвивається відносний гіперінсулінізм, що проявляється зниженням рівня глюкози в крові, пригнічується кількість альбумінів в печінці, РНК, знижується активність внутріклітинних ферментів крові, зменшується адаптація до стресових ситуацій та до інфекцій. Недостатнє утворення андрогенів, поряд з порушенням функції статевих залоз, веде до погіршення анаболітичних процесів. Дегідратація, збіднення м’язів глікогеном, зниження анаболітичних процесів в тканинах і особливо в м’язах, що супроводжується гіпоплазією, атрофією м’язів, а також анорексія ведуть до прогресуючої втрати маси тіла.
При туберкульозній етіології захворювання уражуються два наднирники. Наднирники збільшені, маса їх досягає 60 – 80 г. На зрізі спостерігаються центральні ділянки казеозного некрозу, що містять вапнякові відкладення, оточені грануляційною тканиною з епітеліоїдних чи гігантських клітин, які переходять в шар сполучнотканинної капсули, містять велику кількість лімфоцитів. Тканина наднирників залишається у вигляді острівків, спостерігаються ознаки регенерації і функціональної активності. Важкі ураження наднирників з розростанням сполучної тканини можуть бути при порушеннях кровообігу, інфекційно-токсичних станах.
У випадку первинної деструктивної атрофії в кірковій речовині виявляються великі лімфоцитарні інфільтрати, які замінюються на фіброзну тканину. Між вогнищами ураження є залишки залозистої тканини з ознаками підвищеної функціональної активності. При амілоїдозі відбувається відкладення амілоїда в корі звичайно навколо капілярів пучкової зони, клітини кори стискаються глибами амілоїду і атрофуються. При вторинному гіпокортицизмі в результаті зменшення виділення кортикотропіну відбувається атрофія наднирників, їх зменшення, змінюється будова – дещо розширюються клубочкова і сітчаста зони за рахунок витончення пучкової.
Хронічна недостатність кори наднирників нерідко супроводжується гіпоплазією вилочкової залози і лімфоїдного апарату, лімфоцитарною інфільтрацією і фіброзом щитовидної залози. В деяких випадках спостерігаються поверхневі, різко окреслені виразки слизової оболонки шлунку.
В залежності від вираженості клінічної симптоматики і важкості стану адісонова хвороба має три форми: легку, середню, важку.
Хворі з легкою формою захворювання почувають себе задовільно, зберігають працездатність, не потребують систематичної гормонозамінної терапії. Достатні дієтичні корективи – додаток кухонної солі, обмеження споживання продуктів з великим вмістом калію, а також прийом аскорбінової кислоти. Але будь-яке перевантаження чи стресова ситуація можуть привести до погіршення стану, навіть до розвитку гострої наднирникової недостатності, тому в ці періоди необхідно вводити гормони.
При середній важкості захворювання хворі потребують систематичного прийому невеликих доз гормонів.
При важкій формі навіть короткочасна відміна наднирникових гормонів приводить до погіршення стану і розвитку адісонічного кризу.
Адісонова хвороба може спостерігатися на протязі багатьох років без вираженого погіршення стану. При наявності активного туберкульозного процесу хвороба порівняно прогресує і приводить до інвалідності. Періодично при хронічному стані у зв’язку з стресами, інфекціями стан хворого поступово погіршується, наростає вираженість основних симптомів, чіткіше проявляються судинні і шлунково-кишкові розлади, посилюються адинамія, пігментація, дегідратація. Без вчасної допомоги хворий впадає в коматозний стан.
Адісонічний криз
Важким проявом адісонової хвороби є гостра недостатність кори наднирників, яка при відсутності вчасного лікування закінчується смертю. Причиною розвитку адісонічного кризу можуть бути різноманітні стресові ситуації: інфекції, інтоксикації, психічні і фізичні перевантаження, оперативні втручання, перерва в прийомі ліків при середній і важкій формах адісонової хвороби. Гостра недостатність кори наднирників в деякий випадках є першим проявом стертої форми адісонової хвороби.
Гормональнопродукуючі пухлини наднирників
1. Феохромоцитома – гормонально-активна пухлина, що секретує в підвищеній кількості катехоламіни з локалізацією в мозковій речовині наднирників або в симпатичних гангліях.
В етіології певну роль відіграє спадковість. Описані сімейні випадки захворювання феохромоцитомою.
Патогенез захворювання пов’язаний з поступленням в кров великої кількості адреналіну і норадреналіну, що виробляється пухлиною.
2. Кортикостерома – гормонально-активна пухлина кори наднирників, що виходить з пучкової зони кори і супроводжується гіперпродукцією глюкокортикоїдів. Клінічна картинна кортикостероми тотожна з захворюванням Іценко-Кушинга діенцефально-гіпофізарного генезу. З’являється підвищена втома і м’язева слабкість, ожиріння за диспластичним типом. Пухлина може бути доброякісною і злоякісною. Злоякісні пухлини швидко прогресують.
3. Кортикоестрома - гормонально-активна пухлина кори наднирників (сітчастої та пучкової зон), що характеризується надмірним виділенням естрогенів, іноді і глюкокортикоїдів. Клінічно проявляється синдромом фемінізації і симптомами хвороби Іценко-Кушинга.
4. Андростерома – гормонально-активна пухлина кори наднирників (сітчастої зони), щохарактеризується надмірною продукцією кортикостероїдів (андрогенів).
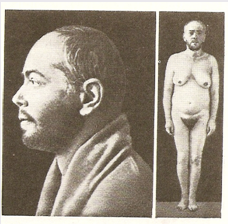
Фото 3. Результат гіперфункції кори наднирників (за В.Г.Барановим)
Андрогенітальний синдром
Захворювання пов’язане з генетичним ураженням ферментних систем, що приймають участь в синтезі глюкокортикоїдів, і, як наслідок цього, - підвищена секреція корою наднирників стероїдних гормонів, частіше андрогенних.
В основі захворювання лежить генетично зумовлена неповноцінність тих чи інших ферментних систем, що приймають участь у біосинтезі стероїдних гормонів. Джерелом утворення гормонів кори наднирників є холостерин, який під впливом ряду ферментів послідовно перетворюється в прегненолон і прогестерон. Останній є вихідним продуктом для синтезу гліко і мінералокортикоїдів.
В результаті дії рецесивного гена може уражатися один з ферментів, що приводить до порушення утворення кортизолу, дефіцит якого опосередкований через гіпоталамус. Безпосередньо через гіпофіз викликає надмірне утворення кортикотропіну, гіперфункцію і гіпертрофію кори наднирників. Різко виростає утворення андрогеенів, синтез їх збільшується, що впливає на розвиток статевих органів. Надмірне утворення андрогенів викликає передчасну ізосексуальну (у хлопчиків) і гетеросексуальну (у дівчаток) сексуальну поведінку, характер якої залежить від ступеню порушення і часу його прояву. Характерна значна гіперплазія кори наднирників, переважно сітчастої зони.
Гостра недостатність кори наднирників (синдром Уотерхауза – Фрідеріксена)
Гостра недостатність кори наднирників – це стан, який характеризується швидким розвитком важких порушень водно-електролітного обміну, гострою нестачею кровообміну, неврологічних та диспептичних явищ в результаті раптового випадіння функції кори наднирників.
Гостра недостатність кори наднирників розвивається у новонароджених, дітей, людей молодого віку.
У новонароджених захворювання може бути викликане крововиливом в кору наднирників в результаті важких пологів, що супроводжувалися асфікцією або накладанням щипців, токсикозом вагітних. Крововилив в кору наднирників можливий при інфекційних захворюваннях: грип, кір, скарлатина, дифтерія, дизентерія, при менінгококовому чи стафілококовому сепсисі, при гемораргічному діатезі, гемофілії та інших захворюваннях крові. Захворювання може розвиватися при тромбозі наднирникових вен, опіках.
Гостра недостатність кори наднирників розвивається при видаленні гормонально-активної пухлини кори наднирників.
В результаті раптового ураження кори наднирників і виникнення дефіциту глюко- і мінералокортикоїдів катастрофічно швидко виникають важкі порушення обміну, властиві для адісонової хвороби, порушується функція різних органів і систем, бурхливо розвивається стан, що нагадує важку форму адісоноового кризу, що частог приводить до смерті.
При гострій недостатністі кори наднирників в результаті порушення кровообігу судини наднирників переповнені кров’ю, інколи відмічається велика кількість крововиливів. Залишки тканини погано диференціюються. Окремі ділянки гемораргій можуть бути у вигляді смужок по ходу судин, звичайно в кірковій речовині.
Висновки
Підводячи підсумки нашої роботи, хочеться відмітити наступні твердження.
1. У чотиримісячних плодів маса наднирників більша за нирки або маси їх однакові. Більшу частину залози в цей період займає кора. У дітей до восьмого року життя кіркова речовина наднирників досягає структурної зрілості і отримує морфологічну стійкість. В такому стані наднирники залишаються до старечого віку. Якщо в період молодості і зрілості особливо інтенсивно розвиваються клубочкова і сітчаста зони, то в процесі старіння ці зони кори наднирників інтенсивніше інволюють, ніж пучкова.
На відміну від інших органів (серце, нирки, легені та ін.), які в утробному періоді не досягають необхідної величини і рівня функціональної активності, не лише наднирники, а й інші ендокринні залози виявляються достатньо великими і зрілими у функціональному відношенні.
2. В процесі старіння відмічаються неоднозначні зміни у функції різних шарів і зон кори наднирників. При фізіологічному протіканні процесів старіння (навіть до пізніх термінів онтогенезу) наднирники є надійною ланкою в здійсненні важливих механізмів адаптації. Вивчення онтогенезу наднирників у ссавців свідчить про те, що ці органи рано володіють здатністю до здійснення секреторної активності і реагують на стреси. Відповідно, вони приймають участь не лише в регуляції росту и розвитку організму, але і в реакціях адаптації. Функціональна активність і резерви наднирників плоду залежать від характеру патологічного впливу, його сили і тривалості, що має важливе значення для розробки методів стимуляції і гормонозамінної терапії при лікуванні плодів та новонароджених.
3. Наднирникові залози можуть мати різноманітні паталогічні стани, що пов’язані перш за все з двостороннім ураженням цих залоз, недостатньою стимуляцією наднирників адренокортикотропного гормона на грунті гіпоталамо-гіпофізарних порушень, різноманітними стресовими ситуаціями: інфекціями, інтоксикаціями, психічними і фізичними перевантаженнями, оперативними втручаннями. Певну роль відіграє спадковість.
Найпоширенішим захворюванням наднирників є адісонова хвороба – хронічна недостатність кори наднирників, що розвивається внаслідок двостороннього ураження цих залоз або недостатньої стимуляції наднирників адренокортикотропного гормона на грунті гіпоталамо-гіпофізарних порушень.
У новонароджених захворювання можуть бути викликані крововиливом в кору наднирників в результаті важких пологів, що супроводжувалися асфікцією або накладанням щипців, токсикозом вагітних. Крововилив в кору наднирників можливий при інфекційних захворюваннях.
Патологічними станами наднирників є гормональнопродукуючі пухлини: феохромоцитома, кортикостерома, кортикоестрома, андроестрома.
Список використаної літератури
Книги, монографії
Один автор
1. Алтухова В.И. Гистохимическое выявление активности дегидрогеназы в коре надпочечников плодов человека. [для студентів вищих навчальних закладів]/ В.И. Алтухова [Гормональные факторы индивидуального развития.] М.: Наука, 1974. – С. 291 – 303.
2. Артишевский А.А. Надпочечные железы (строение, функция, развитие) [для студентів вищих навчальних закладів]/А.А. Артишевский – Минск: Беларусь, 1977. – 126 с.
3. Багрий А.В. Эндокринология. [для студентів вищих навчальних закладів] / А.В. Багрий [Новейший справочник.] – М.: Изд-во Эксмо, 2005. – С.19 – 22.
4. Балаболкин М.И. Эндокринология. [для студентів вищих навчальних закладів]/ М.И. Балаболкин – М.: Медицина, 1989. – С. 109.
5. Дедов И.И. Болезни органов эндокринной системы [для студентів вищих навчальних закладів]/ Под ред. И.И. Дедова– М.: Медицина, 2000. – С. 58.
6. Ефимов А.С. Эндокринология. [для студентів вищих навчальних закладів] / А.С. Ефимов – Киев: Вища школа, 1983. - С.208.
7. Матешук-Вацеба Л.Р. Нормальна анатомія. [для студентів вищих навчальних закладів]/ Л.Р. Матешук-Вацеба– Львів, Поклик сумління, 1997. – 156 – 162.
8. Мицкевич М.С. Гормональные регуляции в онтогенезе животных. / М.С. Мицкевич– М.: Наука, 1978. – 174 с.
9. Окороков А.Н. Диагностика болезней внутренних органов: [Практическое руководство]: В 3 т. – Т.2.- Витебск: Белмедкнига, 1998.
10. Рудзской С.Л. Использование антропометрических показателей и клинической эндокринологии / С.Л. Рудзской // Клиническая эндокринология и эндокринная хирургия. – 2003.
11. Савельєва Г.М. Акушерство, гинекология, эндокринология. [для студентів вищих навчальних закладів]/ Под ред. Г.М. Савельевой – М.: Медицина, 1999. – С.55.
12. Старкова Н.Т. Клиническая эндокринология. [для студентів вищих навчальних закладів]/ Под ред. Н.Т. Старковой– М.: Медицина, 1991. – С. 88 – 96.
Два автори
13. Алтухова В.И., Кузнецова В.М. Количественное и топографическое изучение холестерина и аскорбиновой кислоты в надпочечниках зародышей крыс. [для студентів вищих навчальних закладів] / В.И.Алтухова, В.М. Кузнецова– В кн. Становление эндокринных функций в зародышевом периоде. М.: Наука, 1974. – С. 108 – 117.
14. Клегг П., Клегг А. Гормоны, клетки, организм. [для студентів вищих навчальних закладів]/ П., Клегг А. Клегг– М.: Мир, . [Перевод с английского] 1974. – С. 135 – 150.
15. Травянко Т.Д., Сольский Я.П. Справочник по акушерско-гинекологической эндокринологии. [для студентів вищих навчальних закладів] / Т.Д. Травянко, Я.П. Сольский Киев: Здоровье, 1995.
16. Хадорн Э., Венер Р. Общая зоология. [для студентів вищих навчальних закладів]/ Э., Хадорн, Р. Венер. – М.: Мир, . [Перевод с английского] 1989. – С. 125.
Три автори
17. Баевский Ю.Б., Леонов Б.В., Лукин В.А. Гормональные регуляции раннего ембриогенеза млекопитающих. [для студентів вищих навчальних закладів] / Ю.Б. Баевский Б.В. Леонов В.А. Лукин – Успехи современной биоло., 1973, 75, вып. 2. - С. 265.
18. Соффер Л., Дорфман Р., Гебрилав Л. Надпочечные железы человека. [для студентів вищих навчальних закладів]/ Л.Соффер, Р. Дорфман, Л. Гебрилав. – М.: Медицина, . [Перевод с английского]1966. – 496 с.
Чотири автори
19. Свечникова Н.В., Вержиковская Н.В., Беккер В.И., Мороз Е.В. Железы внутренней секреции в процессе старения./ Н.В Свечникова., Н.В Вержиковская, В.И Беккер., Е.В. Мороз– К.: Изд-во Здоровья, 1983. – С. 67.
Перепечатка материалов без ссылки на наш сайт запрещена