Учебное пособие: Анатомия центральной нервной системы
Учебное пособие: Анатомия центральной нервной системы
СОЦИАЛЬНО-ТЕХНОЛОГИЧЕСКИЙ ИНСТИТУТ МОСКОВСКОГО ГОСУДАРСТВЕННОГО УНИВЕРСИТЕТА СЕРВИСА
АНАТОМИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
(Учебное пособие)
О.О. Якименко
Москва - 2002
Пособие по анатомии нервной системы предназначено для студентов Социально-технологического института факультета психологии. Содержание включает основные вопросы, связанные с морфологической организацией нервной системы. Помимо анатомических данных о структуре нервной системы в работу включены гистологические цитологические характеристики нервной ткани. А также вопросы информации о росте и развитии нервной системы от эмбрионального до позднего постнатального онтогенеза.
Для наглядности излагаемого материала в текст внесены иллюстрации. Для самостоятельной работы студентов дан список учебной и научной литературы, а также анатомических атласов.
Классические научные данные по анатомии нервной системы являются фундаментом для изучения нейрофизиологии мозга. Знание морфологических характеристик нервной системы на каждом этапе онтогенеза необходимо для понимания возрастной динамики поведения и психики человека.
РАЗДЕЛ I. ЦИТОЛОГИЧЕСКИЕ И ГИСТОЛОГИЧЕСКИЕ ХАРАКТЕРИСТИКИ НЕРВНОЙ СИСТЕМЫ
Общий план строения нервной системы
Главная функция нервной системы состоит в быстрой и точной передаче информации, обеспечивая взаимосвязь организма с окружающим миром. Рецепторы реагируют на любые сигналы внешней и внутренней среды, преобразуя их в потоки нервных импульсов, которые поступают в центральную нервную систему. На основе анализа потоков нервных импульсов мозг формирует адекватный ответ.
Вместе с эндокринными железами нервная система регулирует работу всех органов. Эта регуляция осуществляется благодаря тому, что спинной и головной мозг связаны нервами со всеми органами, двусторонними связями. От органов в центральную нервную систему поступают сигналы об их функциональном состоянии, а нервная система в свою очередь посылает сигналы к органам, корректируя их функции и обеспечивая все процессы жизнедеятельности - движение, питание, выделение и другие. Кроме того нервная система обеспечивает координацию деятельности клеток, тканей, органов и систем органов, при этом организм функционирует как единое целое.
Нервная система является материальной основой психических процессов: внимания, памяти, речи, мышления и др., с помощью которых человек не только познает окружающую среду, но и может активно ее изменять.
Таким образом, нервная система- это та часть живой системы, которая специализируется на передаче информации и на интегрировании реакций в ответ на воздействие окружающей среды.
Центральная и периферическая нервная система
Нервная система по топографическому признаку подразделяется на центральную нервную систему, куда входит головной мозг и спинной мозг, и периферическую, которая состоит из нервов и ганглиев.

 Нервная система
Нервная система
| Центральная нервная система | Периферическая нервная система |
![]()
![]()
![]()
![]()
| головной мозг | спинной мозг | ганглии | нервы |
Согласно классификации по функциональному признаку нервная система подразделяется на соматическую (отделы нервной системы, регулирующие работу скелетных мышц) и автономную (вегетативную), которая регулирует работу внутренних органов. В автономной нервной системе выделяют два отдела: симпатический и парасимпатический.
![]()
![]() Нервная система
Нервная система
![]()
![]() соматическая автономная
соматическая автономная
симпатическая парасимпатическая
Как соматическая, так и автономная нервная системы включают центральный и периферический отделы.
Нервная ткань
Основной тканью, из которой образована нервная система, является нервная ткань[1]. Она отличается от других видов ткани тем, что в ней отсутствует межклеточное вещество.
Нервная ткань состоит из двух видов клеток: нейронов и глиальных клеток. Нейроны играют главную роль, обеспечивая все функции центральной нервной системы. Глиальные клетки имеют вспомогательное значение, выполняя опорную, защитную, трофическую функции и др. В среднем количество глиальных клеток превышает количество нейронов в отношении 10:1 соответственно.
Оболочки мозга образованы соединительной тканью, а полости мозга - особым видом эпителиальной ткани (эпиндимная выстилка).
Нейрон - структурно-функциональная единица нервной системы
Нейрон обладает признаками, общими для всех клеток: имеет оболочку-плазматическую мембрану, ядро и цитоплазму. Мембрана представляет собой трехслойную структуру, содержащую липидные и белковые компоненты. Кроме того на поверхности клетки имеется тонкий слой, называемый гликокалисом. Плазматическая мембраны регулирует обмен веществ между клеткой и средой. Для нервной клетки это особенно важно, так как мембрана регулирует движение веществ, которые непосредственно связаны с нервной сигнализацией. Также мембрана служит местом электрической активности, лежащей в основе быстрой нервной сигнализации и местом действия пептидов и гормонов. Наконец, ее участки образуют синапсы - место контакта клеток.
Каждая нервная клетки обладает ядром, которое содержит генетический материал в форме хромосом. Ядро выполняет две важных функции - контролирует дифференцировку клетки в ее конечную форму, определяя виды связей и регулирует синтез белка во всей клетке, управляя ростом и развитием клетки.
В цитоплазме нейрона имеются органеллы (эндоплазматический ретикулум, аппарат Гольджи, митохондрии, лизосомы, рибосомы и др.).
Рибосомы синтезируют белки, часть которых остается в клетке, другая часть предназначена для выведения из клетки. Кроме того, рибосомы производят элементы молекулярного аппарата для большей части клеточных функций: ферменты, белки-переносчики, рецепторы, белки мембран и т.д
Эндоплазматический ретикулум представляет систему каналов и окруженных мембраной пространств (крупных, плоских, называемых цистернами, и мелких, называемых везикулами или пузырьками) Выделяют гладкий и шероховатый эндоплазматический реьтикулом. Последний содержит рибосомы
Функция аппарата Гольджи состоит в хранении, концентрировании и упаковке секреторных белков.
Кроме систем, вырабатываающих и переносящих разные вещества, клетка обладает внутренней пищеварительной системой, состоящей из лизосом, не имеющих определеной формы. Они содержат разнообразные гидролитические ферменты, которые расщепляют и переваривают множество соединений, возникающих как внутри, так и вне клетки.
Митохондрии- это самая сложная органела клетки после ядра. Ее функция - выработка и доставка энергии, нееобходимая для жизнедеятельности клеток.
Большая часть клеток тела способна усваивать различные сахара, при этом энергия или выделяется или запасается в клетке ввиде гликогена. Однако нервные клетки в головном мозгу используют исключительно глюкозу, так как все другие вещества задерживаются гематоэнцефалическим барьером. Большинство из них лишены способности запасать гликоген, что усиливает их зависимость в отношении энергии от глюкозы в крови и от кислорода. Поэтому в нервных клетках самое большое количество митохондрий.
В нейроплазме содержатся органеллы специального назначения: микротрубочки и нейрофиламенты, которые различаются размером и строением. Нейрофиламенты встречаются только в нервных клетках и представляют внутренний скелет нейроплазмы. Микротрубочки тянутся вдоль аксона по внутренним полостям от сомы до окончания аксона. Эти органеллы и распространяют биологически активные вещества (рис. 1 А и Б). Внутрииклеточный транспорт между телом клетки и отходящими от него отростками может быть ретроградным- от нервных окончаний к телу клетки и ортоградным – от тела клетки к окончаниям.
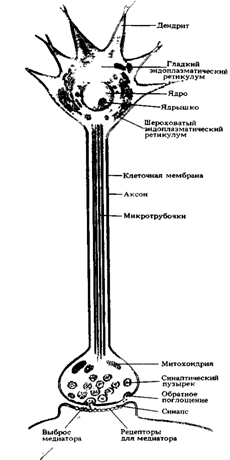
Рис. 1 А. Внутреннее строение нейрона
Отличительной особенностью нейронов является наличие митохондрий в аксоне как добавочного источника энергии и нейрофибрилл. Взрослые нейроны не способны к делению.
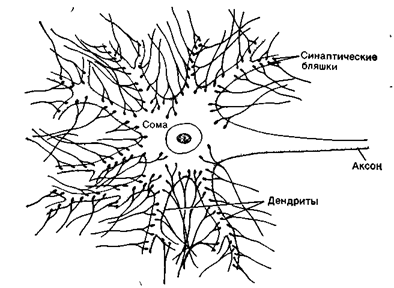
Каждый нейрон имеет расширенную центральную часть тело - сому и отростки- дендриты и аксон. Тело клетки заключено в клеточную оболочку и содержит ядро и ядрышко, поддерживая целостность мембран тела клетки и ее отростков, обеспечивающих проведение ими нервных импульсов. По отношению к отросткам сома выполняет трофическую функцию, регулируя обмен веществ клетки. По дендритам ( афферентные отростки ) импульсы поступают к телу нервной клетки, а по аксонам ( эфферентные отростки ) от тела нервной клетки к другим нейронам или органам
ольшинство дендритов (дендрон - дерево) короткие, сильно ветвящиеся отростки. Их поверхность значительно увеличивается за счет небольших выростов – шипиков. Аксон (аксис - отросток) чаще длинный, мало ветвящийся отросток.
Каждый нейрон имеет только один аксон, длина которого может достигать несколько десятков сантиметров. Иногда от аксона отходят боковые отростки - коллатерали. Окончания аксона, как правило, ветвятся и их называют терминалями. Место, где от сомы клеток отходит аксон, называется аксональным холмиком.
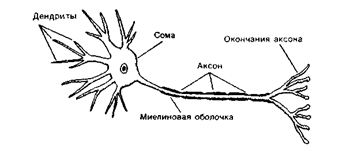
Рис. 1 Б. Внешнее строение нейрона
Существует несколько классификаций нейронов, основанных на разных признаках: форме сомы, количеству отростков, функциям и эффектам, которые нейрон оказывает на другие клетки.
В зависимости от формы сомы различают зернистые (ганглиозные) нейроны, у которых сома имеет округлую форму; пирамидные нейроны разных размеров - большие и малые пирамиды; звездчатые нейроны; веретенообразные нейроны (рис.2 А).
По количеству отростков выделяют униполярные нейроны, имеющие один отросток, отходящий от сомы клеток; псевдоуниполярные нейроны (такие нейроны имеют Т-образный ветвящийся отросток); биполярные нейроны, имеющие один дендрит и один аксон и мультиполярные нейроны, которые имеют несколько дендритов и один аксон (рис. 2 Б).
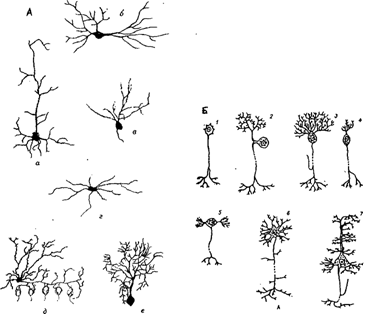
| Различные типы нервных клеток, а - пирамидная, б - двойная пирамидная клетка гиппокампа, в - клетка-зерно зубчатой извилины, г - веретенообразная клетка, д - корзинчатая клетка, е - клетка Пуркинье. | 1 - униполярный нейрон в среднемозговом ядре тройничного нерва; 2 - псевдоуниполярный нейрон спинального ганглия; 3 - грушевидная клетка коры мозжечка; 4 - биполярный нейрон вестибулярного ганглия; 5 -мультиполярный вегетативный нейрон с перпендикулярным расположением аксона и дендритов; 6 - звездчатая клетка; 7 - пирамидная клетка. |
Рис. 2. Классификация нейронов по форме сомы, по количеству отростков
Униполярные нейроны находятся в сенсорных узлах (например, спинальных, тройничном) и связаны с таким видом чувствительности, как болевая, температурная, тактильная, чувством давления, вибрации и т.д.
Эти клетки, хотя и называют униполярными, на самом деле имеют два отростка, которые сливаются вблизи тела клетки.
Биполярные клетки характерны для зрительной, слуховой и обонятельной систем
Мультиполярные клетки имеют разнообразную форму тела – веретенообразную, корзинчатую, звездчатую, пирамидную - малой и большой формы.
По выполняемым функциям нейроны бывают: афферентные, эфферентные и вставочные (контактные).
Афферентные нейроны - сенсорные (псевдоуниполярные), их сомы расположены вне центральной нервной системы в ганглиях (спинномозговых или черепно-мозговых). Форма сомы - зернистая. Афферентные нейроны имеют один дендрит, который подходит к рецепторам (кожи, мышц, сухожилий и т.д.). По дендритам информация о свойствах раздражителей передается на сому нейрона и по аксону в центральную нервную систему.
Эфферентные (двигательные) нейроны регулируют работу эффекторов (мышц, желез, ткани и т.д.). Это мультиполярные нейроны, их сомы имеют звездчатую или пирамидную форму, лежащие в спинном или головном мозге или в ганглиях автономной нервной системы. Короткие, обильно ветвящиеся дендриты воспринимают импульсы от других нейронов, а длинные аксоны выходят за пределы центральной нервной системы и в составе нерва идут к эффекторам (рабочим органам), например, к скелетной мышце.
Вставочные нейроны (интернейроны, контактные) составляют основную массу мозга. Они осуществляют связь между афферентными и эфферентными нейронами, перерабатывают информацию, поступающую от рецепторов в центральную нервную систему. В основном это мультиполярные нейроны звездчатой формы.
Среди вставочных нейронов различаются нейроны с длинными и короткими аксонами (рис. 3 А, Б).
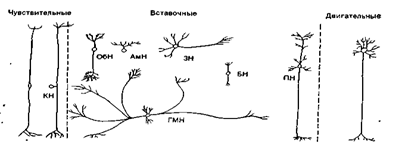
В качестве сенсорных нейронов изображены: нейрон, отросток которого идет в составе слуховых волокон пред-дверно-улиткового нерва (VIII пара), нейрон, реагирующий на стимуляцию кожи (КН). Вставочные нейроны представлены амакрино-вой (АмН) и биполярной (БН) клетками сетчатки, нейроном обонятельной луковицы (ОбН), нейроном голубоватого места (ГМН), пирамидной клеткой коры головного мозга (ПН) и звездчатым нейроном (ЗН) мозжечка. В качестве двигательного нейрона изображен мотонейрон спинного мозга.
Рис. 3 А. Классификация нейронов по выполняемым функциям
|
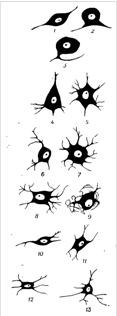
Сенсорные нейрон:
1 - биполярный, 2 - псевдобиполярный, 3 - псевдоуниполярный, 4 - пирамидная клетка, 5 - нейрон спинного мозга, 6 -нейрон п. ambiguus, 7 - нейрон ядра подъязычного нерва. Симпатические нейроны: 8 - из звездчатого ганглия, 9 - из верхнего шейного ганглия, 10 - из интермедиолатеральеного столба бокового рога спинного мозга. Парасимпатические нейроны: 11 - из узла мышечного сплетения кишечной стенки, 12 - из дорсального ядра блуждающего нерва, 13 - из ресничного узла.
По эффекту, который нейроны оказывают на другие клетки, различают возбуждающие нейроны и тормозные нейроны. Возбуждающие нейроны оказывают активизирующий эффект, повышая возбудимость клеток, с которыми они связаны. Тормозные нейроны, напротив, снижают возбудимость клеток, вызывая угнетающий эффект.
Глия
Пространство между нейронами заполнено клетками, которые называются нейроглией (термин глия- обозначает клей, клетки “склеивают” компоненты ЦНС в единое целое). В отличие от нейронов клетки нейроглии делятся в течение всей жизни человека. Нейроглиальных клеток очень много; в некоторых отделах нервной системы их в 10 раз больше, чем нервных клеток. Выделяют клетки макроглии и клетки микроглии (рис.4).
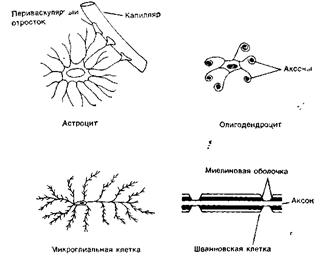
Четыре основных вида клеток глии.
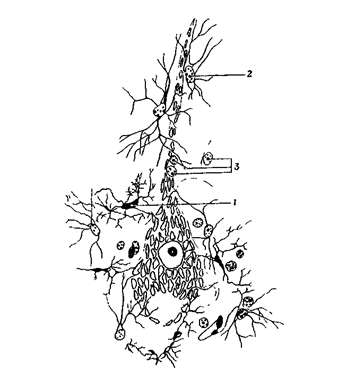
Нейрон, окруженный различными элементами глии
1 - астроциты макроглия
2 - олигодендроциты макроглия
3 – микроглия макроглия
Рис. 4. Клетки макроглии и микроглии
К макроглие относят астроциты и олигодендроциты. Астроциты обладают множеством отростков, которые расходятся от тела клетки во всех направлениях, придавая вид звезды. В центральной нервной системе некоторые отростки заканчиваются концевой ножкой на поверхности кровеносных сосудов. Астроциты, лежащие в белом веществе головного мозга, называются фиброзными астроцитами из-за наличия множества фибрилл в цитоплазме их тел и ветвей. В сером веществе астроциты содержат меньше фибрилл и называются протоплазматическими астроцитами. Они служат опорой нервных клеток, обеспечивают репарацию нервов после повреждения, изолируют и объединяют нервные волокна и окончания, участвуют в метаболических процессах, моделирующих ионный состав, медиаторы. Теперь отвергнуты предположения ь, что они участвуют в транспорте веществ от кровеносных сосудов к нервным клеткам и образуют часть гематоэцефалического барьера.
1. Олигодендроциты меньше по размерам, чем астроциты, содержат небольшие ядра, чаще встречаются в белом веществе и ответственны за формирование миелиновых оболочек вокруг длинных аксонов. Они выполняют роль изолятора и увеличивают скорость проведения нервных импульсов вдоль отростков. Миелиновая оболочка сегментарна, пространство между сегментами называется перехват Ранвье (рис.5). Каждый ее сегмент, как правило, образован одним олигодендроцитом (Шванновская клетка), который, истончаясь, закручивается вокруг аксона. Миелиновая оболочка имеет белый цвет (белое вещество), так как в состав мембран олигодендроцитов входит жироподобное вещество - миелин. Иногда одна глиальная клетка, образуя выросты, принимает участие в образовании сегментов нескольких отростков. Предполагается, что олигодендроциты осуществляют сложный метаболический обмен с нервными клетками.
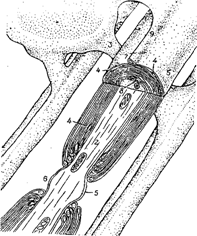
1 - олигодендроцит, 2 - связь между телом клетки глии и миелиновой оболочкой, 4 - цитоплазма, 5 -плазматическая мембрана, 6 - перехват Ранвье, 7 - петля плазматической мембраны, 8 - мезаксон, 9 – гребешок
Рис. 5А. Участие олигодендроцита в образовании миелиновой оболочки
Представлены четыре стадии "обволакивания" аксона (2) шванновской клеткой (1) и его обертывания несколькими сдвоенными слоями мембраны, которые после сжатия образуют плотную миелиновую оболочку.
Рис. 5 Б. Схема оброзования миелиновой оболочки.
Сома нейрона и дендриты покрыты тонкими оболочками, которые не образуют миелин и составляют серое вещесство.
2. Микроглия представлена мелкими клетками, способными к амебовидному передвижению. Функция микроглии - защита нейронов от воспалений и инфекций (по механизму фагоцитоза - захватывание и переваривание генетически чужеродных веществ). Клетки микроглии доставляют нейронам кислород и глюкозу. Кроме того, они входят в состав гематоэнцефалического барьера, который образован ими и эндотелиальными клетками, образующими стенки кровеносных капилляров. Гематоэнцефалический барьер задерживает макромолекулы, ограничивая их доступ к нейронам.
Нервные волокна и нервы
Длинные отростки нервных клеток называют нервными волокнами. По ним нервные импульсы могут передаваться на большие расстояния до 1 метра.
Классификация нервных волокон основана на морфологических и функциональных признаках.
Нервные волокна, имеющие миелиновую оболочку, называются миелинизированными (мякотными), а волокна, не имеющие миелиновой оболочки - немиелинизированными (безмякотными).
По функциональным признакам различают афферентные (чувствительные) и эфферентные (двигательные) нервные волокна.
Нервные волокна, выходящие за пределы нервной системы, образуют нервы. Нерв - это совокупность нервных волокон. Каждый нерв имеет оболочку и кровоснабжение (рис.6).
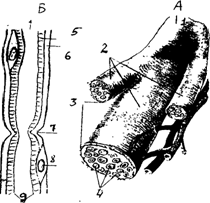
1 - общий ствол нерва, 2 - разветвления нервного волокна, 3 - оболочка нерва, 4 - пучки нервных волокон, 5 - миелиновая оболочка, 6 - мембрана швановской клетки, 7 - перехват Ранвье, 8 - ядро швановской клетки, 9 - аксолемма.
Рис. 6 Строение нерва (А) и нервного волокна (Б).
Различают спинномозговые нервы, связанные со спинным мозгом (31 пара) и черепно-мозговые нервы (12 пар), связанные с головным мозгом. В зависимости от количественного соотношения афферентных и эфферентных волокон в составе одного нерва различают чувствительные, двигательные и смешанные нервы. В чувствительных нервах преобладают афферентные волокна, в двигательных - эфферентные, в смешанных - количественное соотношение афферентных и эфферентных волокон приблизительно равно. Все спинномозговые нервы являются смешанными нервами. Среди черепно-мозговых нервов выделяют три вышеперечисленных типа нервов. I пара - обонятельные нервы (чувствительные), II пара - зрительные нервы (чувствительные), III пара - глазодвигательные (двигательные), IV пара - блоковые нервы (двигательные), V пара - тройничные нервы (смешанные), VI пара - отводящие нервы (двигательные), VII пара - лицевые нервы (смешанные), VIII пара - вестибуло-кохлеарные нервы (смешанные), IX пара - языкоглоточные нервы (смешанные), X пара - блуждающие нервы (смешанные), XI пара - добавочные нервы (двигательные), XII пара - подъязычные нервы (двигательные) (рис 7 ).
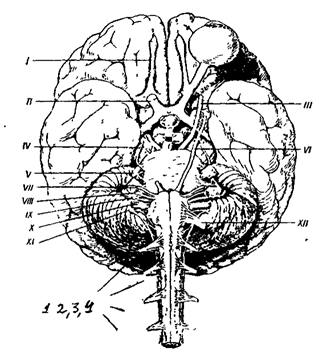
I - пара- обонятельные нервы,
II - пара- зрительные нервы,
III - пара- глазодвигательные нервы,
IV - пара- блоковые нервы,
V - пара - тройничные нервы,
VI - пара- отводящие нервы,
VII - пара- лицевые нервы,
VIII - пара- кохлеарные нервы,
IX - пара- языкоглоточные нервы,
X - пара - блуждающие нервы,
XI - пара- добавочные нервы,
XII - пара-1,2,3,4 - корешки верхних спиномозговых нервов.
Рис. 7, Схема расположения черепно-мозговых и спинальных нервов
Серое и белое вещество нервной системы
На свежих срезах мозга видно, что одни структуры более темные - это серое вещество нервной системы, а другие структуры более светлые - белое вещество нервной системы. Белое вещество нервной системы образовано миелинизированными нервными волокнами, серое - немиелинизированными частями нейрона - сомами и дендритами.
Белое вещество нервной системы представлено центральными трактами и периферическими нервами. Функция белого вещества - передача информации от рецепторов в центральную нервную систему и от одних отделов нервной системы к другим.
Серое вещество центральной нервной системы образовано корой мозжечка и корой полушарий большого мозга, ядрами, ганглиями и некоторыми нервами.
Ядра
Ядра - скопления серого вещества в толще белого вещества. Они расположены в разных отделах центральной нервной системы: в белом веществе больших полушарий - подкорковые ядра, в белом веществе мозжечка - мозжечковые ядра, некоторые ядра расположены в промежуточном, среднем и продолговатом мозге. Большинство ядер являются нервными центрами, регулирующими ту или иную функцию организма.
Ганглии
Ганглии - это скопление нейронов, расположенных вне пределов центральной нервной системы. Различают спинномозговые, черепно-мозговые ганглии и ганглии автономной нервной системы. Ганглии образованы преимущественно афферентными нейронами, но в их состав могут входить вставочные и эфферентные нейроны.
Взаимодействие нейронов
Место функционального взаимодействия или контакта двух клеток (место, где одна клетка оказывает влияние на другую клетку) английский физиолог Ч. Шеррингтон назвал синапсом.
Синапсы бывают периферическими и центральными. Примером периферического синапса является нервно-мышечный синапс, когда нейрон образует контакт с мышечным волокном. Синапсы в нервной системе называются центральными, когда контактируют два нейрона. Выделяется пять типов синапсов, в зависимости от того, какими частями контактируют нейроны: 1) аксо-дендритный (аксон одной клетки контактирует с дендритом другой); 2) аксо-соматический (аксон одной клетки контактирует с сомой другой клетки); 3) аксо-аксональный (аксон одной клетки контактирует с аксоном другой клетки); 4) дендро-дендритный (дендрит одной клетки контактирует с дендритом другой клетки); 5) сомо-соматический (контактируют сомы двух клеток). Основная масса контактов - аксо-дендритных и аксо-соматических.
Синаптические контакты могут быть между двумя возбудительными нейронами, двумя тормозными нейронами или между возбудительным и тормозным нейронами. При этом нейроны, которые оказывают воздействие, называют пресинаптическими, а нейроны, на которые оказывается воздействие - постсинаптическими. Пресинаптический возбудительный нейрон повышает возбудимость постсинаптического нейрона. В этом случае синапс называют возбудительным. Пресинаптический тормозный нейрон оказывает противоположное действие - снижает возбудимость постсинаптического нейрона. Такой синапс называют тормозным. Каждый из пяти типов центральных синапсов имеет свои морфологические особенности, хотя общая схема их строения одинакова.
Строение синапса
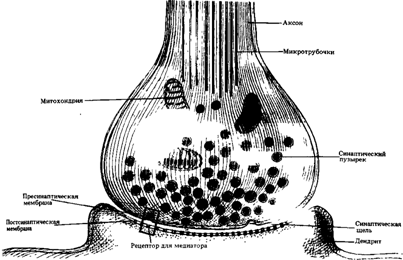
Рассмотрим строение синапса на примере аксо-соматического. Синапс состоит из трех частей: пресинаптического окончания, синаптической щели и постсинаптической мембраны (рис.8 А, Б).
А-Синаптические входы нейрона. Синаптические бляшки окончаний пресинаптичесиих аксонов образуют соединения на дендритах и теле (соме)- постсинаптического нейрона.
Рис. 8 А. Строение синапсов
Пресинаптическое окончание представляет собой расширенную часть терминали аксона. Синаптическая щель - это пространство между двумя контактирующими нейронами. Диаметр синаптической щели составляет 10-20 нм. Мембрана пресинаптического окончания, обращенная к синаптической щели, называется пресинаптической мембраной. Третья часть синапса - постсинаптическая мембрана, которая расположена напротив пресинаптической мембраны.
Пресинаптическое окончание заполнено пузырьками (везикулами) и митохондриями. В везикулах находятся биологически активные вещества - медиаторы. Медиаторы синтезируются в соме и по микротрубочкам транспортируются в пресинаптическое окончание. Наиболее часто в качестве медиатора выступают адреналин, норадреналин, ацетилхолин, серотонин, гамма-аминомасляная кислота (ГАМК), глицин и другие. Обычно синапс содержит один из медиаторов в большем количестве по сравнению с другими медиаторами. По типу медиатора принято обозначать синапсы: адреноэргические, холинэргические, серотонинэргические и др.
В состав постсинаптической мембраны входят особые белковые молекулы - рецепторы, которые могут присоединить молекулы медиаторов.
Синаптическая щель заполнена межклеточной жидкостью, в которой находятся ферменты, способствующие разрушению медиаторов.
На одном постсинаптическом нейроне может находиться до 20000 синапсов, часть из которых являются возбудительными, а часть - тормозными (рис.8 Б).
Б. Схема выброски медиатора и процессов, происходящих в гипотетическом центральном синапсе.
Рис. 8 Б. Строение синапсов
Помимо химических синапсов, в которых при взаимодействии нейронов участвуют медиаторы, в нервной системе встречаются электрические синапсы. В электрических синапсах взаимодействие двух нейронов осуществляется посредством биотоков. В центральной нервной системе преобладают химические стимулы.
В некоторых межнейронах синапсах электрическая и химическая передача осуществляется одновременно - это смешанный тип синапсов.
Влияние возбудительных и тормозных синапсов на возбудимость постсинаптического нейрона суммируется и эффект зависит от места расположения синапса. Чем ближе синапсы расположены к аксональному холмику, тем они эффективнее. Напротив, чем дальше расположены синапсы от аксонального холмика (например, на окончании дендритов), тем они менее эффективны. Таким образом, синапсы, расположенные на соме и аксональном холмике, оказывают влияние на возбудимость нейрона быстро и эффективно, а влияние удаленных синапсов медленно и плавно.
Нейронные сети
Благодаря синаптическим связям нейроны объединены в функциональные единицы - нейронные сети. Нейронные сети могут быть образованы нейронами, расположенными на небольшом расстоянии. Такую нейронную сеть называют локальной. Кроме того, в сеть могут быть объединены нейроны, удаленные друг от друга, из разных областей мозга. Самый высокий уровень организации связей нейронов отражает соединение нескольких областей центральной нервной системы. Такую нервную сеть называют путем или системой. Различают нисходящие и восходящие пути. По восходящим путям информация передается от нижележащих областей мозга к вышележащим (например, от спинного мозга к коре полушарий большого мозга). Нисходящие пути связывают кору больших полушарий мозга со спинным мозгом.
Самые сложные сети называются распределительными системами. Они образуются нейронами разных отделов мозга, управляющих поведением, в которых участвует организм как единое целое.
Некоторые нервные сети обеспечивают конвергенцию (схождение) импульсов на ограниченном количестве нейронов. Нервные сети могут быть построены также по типу дивергенции (расхождение). Такие сети обуславливают передачу информации на значительные расстояния. Кроме того, нервные сети обеспечивают интеграцию (суммирование или обобщение) различного рода информации (рис.9).
|
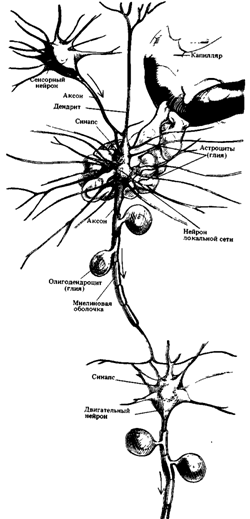
Рис. 9. Нервная ткань.
Крупный нейрон с множеством дендритов получает информацию через синаптический контакт с другим нейроном (в левом верхнем углу). С помощью миелинизированного аксона образуется синаптический контакт с третьим нейроном (внизу). Поверхности нейронов изображены без клеток глии, которые окружают отросток, направленный к капилляру (справа вверху).
Рефлекс как основной принцип работы нервной системы
Одним из примеров нервных сетей может быть рефлекторная дуга, необходимая для осуществления рефлекса. И.М. Сеченов в 1863 г. в работе “Рефлексы головного мозга” развил представление о том, что рефлекс является основным принципом работы не только спинного, но и головного мозга.
Рефлекс - это ответная реакция организма на раздражение при участии центральной нервной системы. Для каждого рефлекса имеется своя рефлекторная дуга - путь, по которому возбуждение проходит от рецептора до эффектора (исполнительного органа). В состав любой рефлекторной дуги входят пять составных частей: 1) рецептор - специализированная клетка, предназначенная для восприятия раздражителя (звуковой, световой, химический и т.д.), 2) афферентный путь, который представлен афферентными нейронами, 3) участок центральной нервной системы, представленный спинным или головным мозгом; 4) эфферентный путь состоит из аксонов эфферентных нейронов, выходящих за пределы центральной нервной системы; 5) эффектор - рабочий орган (мышца или железа и т.д.).
Простейшая рефлекторная дуга включает два нейрона и называется моносинаптической (по числу синапсов). Более сложная рефлекторная дуга представлена тремя нейронами (афферентным, вставочным и эфферентным) и называется трехнейронной или дисинаптической. Однако большинство рефлекторных дуг включает большое количество вставочных нейронов, и называются полисинаптическими (рис. 10 А, Б ).
Рефлекторные дуги могут проходить только через спинной мозг (отдергивание руки при прикосновении к горячему предмету) или только головной мозг (закрывание век при струе воздуха, направленной в лицо), или как через спинной, так и через головной мозг.
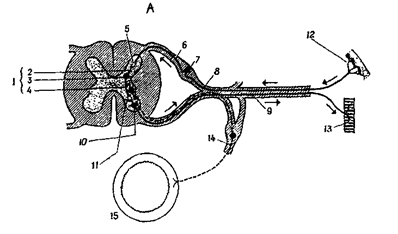
Рис. 10А. 1 - вставочный нейрон; 2 - дендрит; 3 - тело нейрона; 4 - аксон; 5 -синапс между чувствительным и вставочным нейронами; 6 - аксон чувствительного нейрона; 7 - тело чувствительного нейрона; 8 - аксон чувствительного нейрона; 9 - аксон двигательного нейрона; 10 - тело двигательного нейрона; 11 - синапс между вставочным и двигательными нейронами; 12 - рецептор в коже; 13 - мышца; 14 - симпатический гаглий; 15 - кишка.
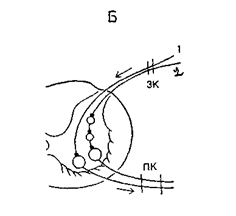
Рис. 10Б. 1 - моносинаптическая рефлекторная дуга, 2 - полисинаптическая рефлекторная дуга, 3К - задний корешок спинного мозга, ПК - передний корешок спинного мозга.
Рис. 10. Схема строения рефлекторной дуги
Рефлекторные дуги замыкаются в рефлекторные кольца с помощью обратных связей. Понятие обратной связи и ее функциональная роль были указаны Беллом в 1826 г. Белл писал, что между мышцей и центральной нервной системой устанавливаются двусторонние связи. С помощью обратной связи в центральную нервную систему поступают сигналы о функциональном состоянии эффектора.
Морфологической основой обратной связи являются рецепторы, расположенные в эффекторе, и афферентные нейроны, связанные с ними. Благодаря обратным афферентным связям осуществляется тонкая регуляция работы эффектора и адекватная реакция организма на изменения окружающей среды.
Оболочки мозга
Центральная нервная система (спинной и головной мозг) имеют три соединительно-тканные оболочки: твердую, паутинную и мягкую. Самая наружная из них твердая мозговая оболочка (она срастается с надкостницей, выстилающей поверхность черепа). Паутинная оболочка лежит под твердой оболочкой. Она плотно прижата к твердой и между ними нет свободного пространства.
Непосредственно к поверхности мозга примыкает мягкая мозговая оболочка, в которой много кровеносных сосудов, питающих мозг. Между паутинной и мягкой оболочками имеется пространство, заполненное жидкостью - ликвором. По составу ликвор близок к плазме крови и межклеточной жидкости и играет противоударную роль. Кроме того, в ликворе содержатся лимфоциты, обеспечивающие защиту от чужеродных веществ. Он же участвует в обмене веществ между клетками спинного, головного мозга и кровью (рис.11 А ).
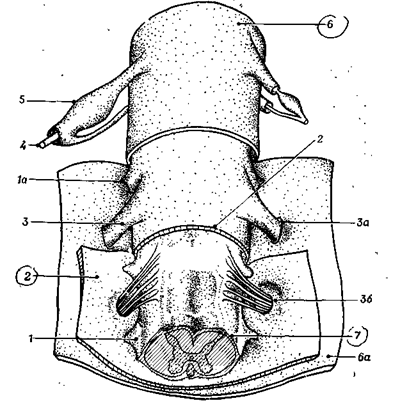
1 - зубчатая связка, отросток которой проходит через расположенную сбоку паутинную оболочку, 1а - зубчатая связка, прикрепленная к твердой мозговой оболочке спинного мозга, 2 - паутинная оболочка, 3 - задний корешок, проходящий в канале, образованном мягкой и паутинной оболочками, За - задний корешок, проходящий через отверстие в твердой мозговой оболочке спинного мозга, 36 - дорсальные ветви спинномозгового нерва, проходящие через паутинную оболочку, 4 - спинномозговой нерв, 5 -спинномозговой узел, 6 - твердая оболочка спинного мозга, 6а - твердая мозговая оболочка, отвернутая в сторону, 7 - мягкая оболочка спинного мозга с задней спинномозговой артерией.
Рис. 11А. Оболочки спинного мозга
Полости мозга
Внутри спинного мозга располагается спинномозговой канал, который, переходя в головной мозг, расширяется в продолговатом мозге и образует четвертый желудочек. На уровне среднего мозга желудочек переходит в узкий канал - Сильвиев водопровод. В промежуточном мозге Сильвиев водопровод расширяется, образуя полость третьего желудочка, который плавно переходит на уровне полушарий головного мозга в боковые желудочки (I и II). Все перечисленные полости также заполнены ликвором ( рис. 11 Б)
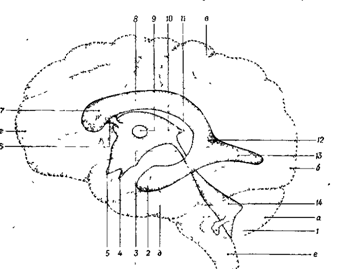
Рис 11Б. Схема желудочков мозга и их отношение к поверхностным структурам полушарий головного мозга.
а - мозжечок, б - затылочный полюс, в - теменной полюс, г - лобный полюс, д - височный полюс, е - продолговатый мозг.
1 - боковое отверстие четвертого желудочка (отверстие Люшка), 2 - нижний рог бокового желудочка, 3 - водопровод, 4 - recessus infundibularis, 5 - recrssus opticus, 6 - межжелудочковое отверстие, 7 - передний рог бокового желудочка, 8 - центральная часть бокового желудочка, 9 - сращение зрительных бугров (massa inter-melia), 10 - третий желудочек, 11 -recessus pinealis, 12 - вход в боковой желудочек, 13 - задний pro бокового желудочка, 14 - четвертый желудочек.
Рис. 11. Оболочки (А) и полости мозга (Б)
РАЗДЕЛ II. СТРОЕНИЕ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Спинной мозг
Внешнее строение спинного мозга
Спинной мозг представляет собой уплощенный тяж, расположенный в позвоночном канале. В зависимости от параметров тела человека его длина составляет 41-45 см, средний диаметр 0.48-0.84 см, вес около 28-32 г. В центре спинного мозга проходит спинномозговой канал, заполненный ликвором, а передней и задней продольными бороздами он поделен на правую и левую половины.
Спереди спинной мозг переходит в головной мозг, а сзади заканчивается мозговым конусом на уровне 2-го позвонка поясничного отдела позвоночника. От мозгового конуса отходит соединительно-тканная концевая нить (продолжение концевых оболочек), которая прикрепляет спинной мозг к копчику. Концевая нить окружена нервными волокнами (конский хвост) (рис. 12).
На спинном мозге выделяется два утолщения - шейное и поясничное, от которых отходят нервы, иннервирующие, соответственно, скелетные мышцы рук и ног.
В спинном мозге выделяют шейный, грудной, поясничный и крестцовый отделы, каждый из которых подразделяется на сегменты: шейный - 8 сегментов, грудной - 12, поясничный - 5, крестцовый 5-6 и 1 - копчиковый. Таким образом, общее количество сегментов - 31 (рис. 13). Каждый сегмент спинного мозга имеет парные спинномозговые корешки - передние и задние. По задним корешкам в спинной мозг поступает информация от рецепторов кожи, мышц, сухожилий, связок, суставов, поэтому задние корешки называют сенсорными (чувствительными). Перерезка задних корешков выключает тактильную чувствительность, но не приводит к утрате движений.
|
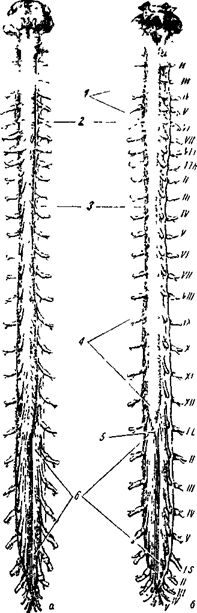
a - вид спереди (вентральная его поверхность);
б - вид сзади (дорсальная его поверхность).
Твердая и паутинная оболочки разрезаны. Сосудистая оболочка снята. Римскими цифрами обозначен порядок расположения шейных (с), грудных (th), поясничных (t)
и крестцовых (s) спинномозговых нервов.
1 - шейное утолщение
2 - спинальный ганглий
3 - твердая оболочка
4 - поясничное утолщение
5 - мозговой конус
6 - концевая нить
Рис. 13. Спинной мозг и спинномозговые нервы (31 пара).
По передним корешкам спинного мозга нервные импульсы поступают к скелетным мышцам тела (за исключением мышц головы), вызывая их сокращение, поэтому передние корешки называют двигательными или моторными. После перерезки передних корешков с одной стороны наблюдается полное выключение двигательных реакций, чувствительность к прикосновению или давлению при этом сохраняется.
Передние и задние корешки каждой стороны спинного мозга объединяются в спинномозговые нервы. Спинномозговые нервы называют сегментарными, их количество соответствует числу сегментов и составляет 31 пару ( рис. 14)
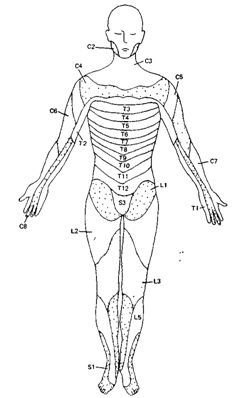
Распределение зон спинномозговых нервов по сегментам было установлено путем определения размеров и границ участков кожи (дерматомов), иннервируемых каждым нервом. Дерматомы расположены на поверхности тела по сегментарному принципу. К шейным дерматомам относятся задняя поверхность головы, шея, плечи и передняя поверхность предплечий. Грудные сенсорные нейроны иннервируют оставшуюся поверхность предплечья, грудь и большую часть живота. Сенсорные волокна поясничных, крестцовых и копчиковых сегментов подходят к остальной части живота и ногам.
Рис. 14. Схема дерматомов. Иннервация поверхности тела 31 парой спинномозговых нервов (С - шейные, Т - грудные, L - поясничные, S - крестцовые).
Внутреннее строение спинного мозга
Спинной мозг построен по ядерному типу. Вокруг спинномозгового канала расположено серое вещество, на периферии - белое. Серое вещество образовано сомами нейронов и ветвящимися дендритами, не имеющими миелиновых оболочек. Белое вещество - это совокупность нервных волокон, покрытых миелиновыми оболочками.
В сером веществе различают передние и задние рога, между которыми лежит межуточная зона. В грудном и поясничном отделах спинного мозга имеются боковые рога.
Серое вещество спинного мозга образовано двумя группами нейронов: эфферентными и вставочными. Основную массу серого вещества составляют вставочные нейроны (до 97%) и только 3% составляют эфферентные нейроны или мотонейроны. Мотонейроны расположены в передних рогах спинного мозга. Среди них различают a- и g-мотонейроны: a-мотонейроны иннервируют волокна скелетных мышц и представляют собой крупные клетки с относительно длинными дендритами; g-мотонейроны представлены мелкими клетками и иннервируют рецепторы мышц, повышая их возбудимость.
Вставочные нейроны участвуют в переработке информации, обеспечивая согласованную работу сенсорных и двигательных нейронов, а также связывают правую и левую половины спинного мозга и его различные сегменты ( рис. 15 А,Б,В )
|
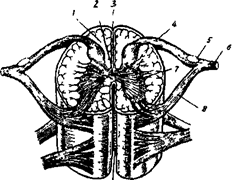
Рис. 15А. 1 - белое вещество мозга; 2 - спинномозговой канал; 3 - задняя продольная борозда; 4 - задний корешок спинномозгового нерва; 5 –спинно-мозговой узел; 6 - спинномозговой нерв; 7 -серое вещество мозга; 8 - передний корешок спинномозгового нерва; 9 - передняя продольная борозда
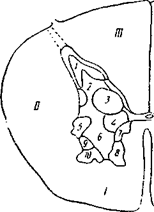
Рис. 15Б. Ядра серого вещества в грудном отделе
1,2,3- чувствительные ядра заднего рога; 4, 5 - вставочные ядра бокового рога; 6,7, 8,9,10 - двигательные ядра переднего рога; I, II, III - передний, боковой и задний канатики белого вещества.
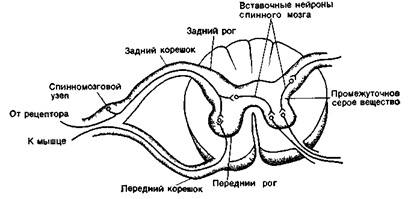
Изображены контакты между чувствительными, вставочными и двигательными нейронами в сером веществе спинного мозга.
Рис. 15. Поперечный разрез спинного мозга
Проводящие пути спинного мозга
Белое вещество спинного мозга окружает серое вещество и образует столбы спинного мозга. Различают передние, задние и боковые столбы. Столбы - это тракты спинного мозга, образованные длинными аксонами нейронов, идущими вверх по направлению к головному мозгу (восходящие пути) либо вниз - от головного мозга к ниже расположенным сегментам спинного мозга (нисходящие пути).
По восходящим путям спинного мозга передается информация от рецепторов мышц, сухожилий, связок, суставов и кожи к головному мозгу. Восходящие пути являются также проводниками температурной и болевой чувствительности. Все восходящие пути перекрещиваются на уровне спинного (или головного) мозга. Таким образом, левая половина головного мозга (кора полушарий и мозжечок) получают информацию от рецепторов правой половины тела и наоборот.
Основные восходящие пути: от механорецепторов кожи и рецепторов опорно-двигательного аппарата - это мышцы, сухожилия, связки, суставы - пучки Голля и Бурдаха или соответственно они же - нежный и клиновидный пучки представлены задними столбами спинного мозга.
От этих же рецепторов информация поступает в мозжечок по двум путям, представленным боковыми столбами, которые называются передним и задним спиномозжечковыми трактами. Кроме того, в боковых столбах проходят еще два пути - это боковой и передний спинно-таламические пути, передающие информацию от рецепторов температурной и болевой чувствительности.
Задние столбы обеспечивают более быстрое проведение информации о локализации раздражений, чем боковой и передний спинно-таламические пути (рис. 16 А).
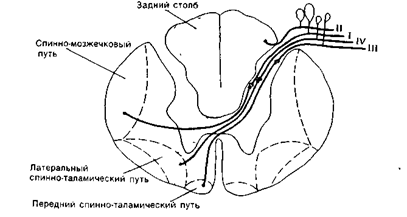
1 - пучок Голля, 2 - пучок Бурдаха, 3 - дорсальный спинно-мозжечковый тракт, 4 - вентральный спинно-мозжечковый тракт. Нейроны группы I-IV.
Рис. 16А. Восходящие пути спинного мозга
Нисходящие пути, проходя в составе передних и боковых столбов спинного мозга, являются двигательными, так как они влияют на функциональное состояние скелетных мышц тела. Пирамидный путь начинается, в основном, в двигательной коре полушарий и проходит к продолговатому мозгу, где большая часть волокон перекрещивается и переходит на противоположную сторону. После этого пирамидный путь разделяется на боковой и передний пучки: соответственно, передний и боковой пирамидные пути. Большинство волокон пирамидных путей оканчивается на вставочных нейронах, а около 20% образуют синапсы на мотонейронах. Пирамидное влияние является возбуждающим. Ретикуло-спинальный путь, руброспинальный путь и вестибулоспинальный путь (экстрапирамидная система) начинаются соответственно от ядер ретикулярной формации, ствола мозга, красных ядер среднего мозга ив вестибулярных ядер продолговатого мозга. Эти пути проходят в боковых столбах спинного мозга, участвуют в координации движений и обеспечении мышечного тонуса. Экстрапирамидные пути, так же как и пирамидные, являются перекрещенными (рис. 16 Б ).
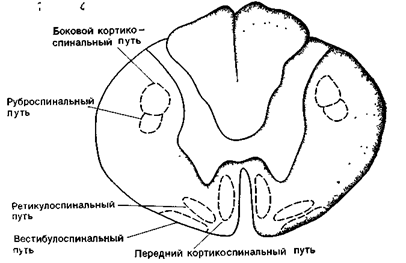
Главные нисходящие спинномозговые пути пирамидной (латеральный и передний кортикоспинальные пути) и экстра пирамидной (руброспинальный, ретикулоспинальный и вестибулоспинальный пути) систем.
Рис. 16 Б. Схема проводящих путей
Таким образом, спинной мозг осуществляет две важнейшие функции: рефлекторную и проводниковую. Рефлекторная функция осуществляется за счет двигательных центров спинного мозга: мотонейроны передних рогов обеспечивают работу скелетных мышц туловища. При этом поддерживается сохранение мышечного тонуса, координации работы мышц сгибателей-разгибателей, лежащих в основе движений и сохранение постоянства позы тела и его частей(рис. 17 А,Б,В ). Мотонейроны, расположенные в боковых рогах грудных сегментов спинного мозга, обеспечивают дыхательными движениями (вдох-выдох, регулируя работу межреберных мышц). Мотонейроны боковых рогов поясничного и крестцового сегментов представляют двигательные центры гладких мышц, входящих в состав внутренних органов. Это центры мочеиспускания, дефекации, работы половых органов.
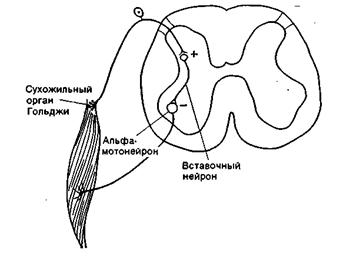
Рис. 17А. Дуга сухожильного рефлекса.
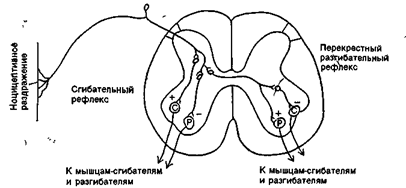
Рис. 17Б. Дуги сгибательного и перекрестного разгибательного рефлекса.
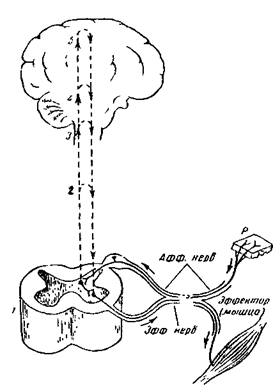
Рис. 17В. Элементарная схема безусловного рефлекса.
Нервные импульсы, возникающие при раздражении рецептора (р) по афферентным волокнам (афф. нерв, показано лишь одно такое волокно) идут к спинальному мозгу (1), где через вставочный нейрон передаются на эфферентные волокна (эфф. нерв), по которым доходят до эффектора. Пунктирные линии - распространение возбуждения от низших отделов центральной нервной системы на ее вышерасположенные отделы (2, 3,4) до коры мозга (5) включительно. Наступающее вследствие этого изменение состояния высших отделов мозга в свою очередь воздействует (см. стрелки) на эфферентный нейрон, влияя на конечный результат рефлекторного ответа.
Рис. 17. Рефлекторная функция спинного мозга
Проводниковую функцию выполняют спинномозговые тракты (рис. 18 А,Б,В,Г,Д ).
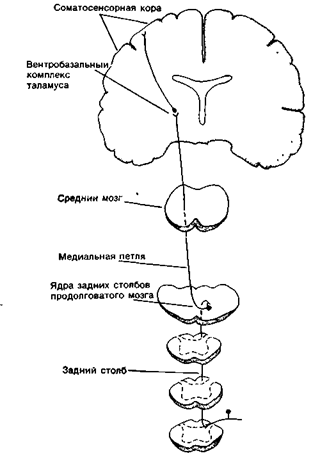
Рис. 18А. Задние столбы. Это цепь, образованная тремя нейронами, передает информацию от рецепторов давления и прикосновения к соматосенсорной коре.
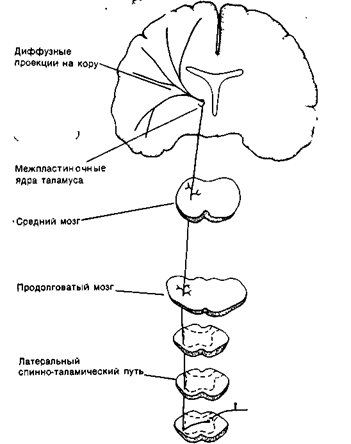
Рис. 18Б. Латеральный спинно-таламический тракт. По этому пути информация от температурных и болевых рецепторов поступает к обширным областям корытоловного мозга.
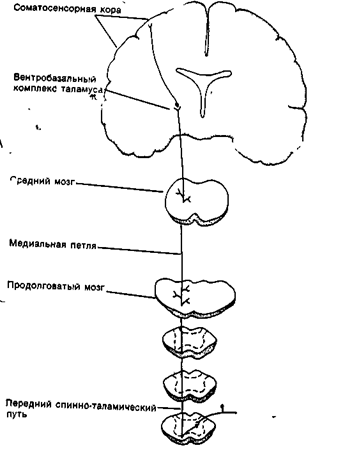
Рис. 18В. Передний спинно-таламический тракт. По этому пути в соматосенсорную кору поступает информация от рецепторов давления и прикосновения, а также от болевых и температурных.
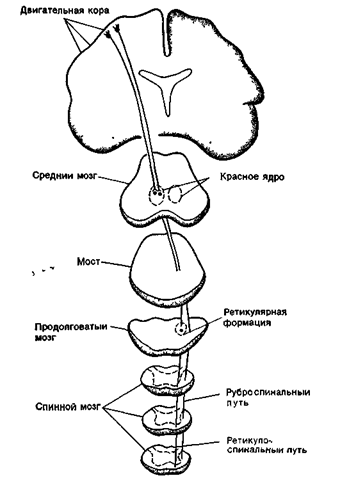
Рис. 18Г. Экстрапирамидная система. Руброспинальный и ретикулоспинальный пути, входящие в состав мультинейронного экстрапирамидного пути, идущего от коры больших полушарий к спинному мозгу.
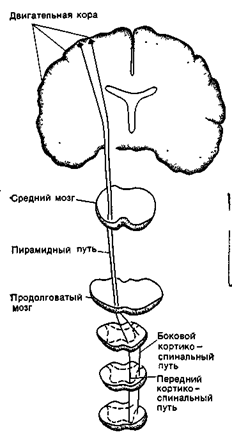
Рис. 18Д. Пирамидный, или кортикоспинальный, путь
Рис. 18. Проводниковая функция спинного мозга
РАЗДЕЛ III. ГОЛОВНОЙ МОЗГ.
Общая схема строения головного мозга ( рис. 19)



 Головной мозг
Головной мозг
|
Задний мозг2 |
Сред-ний мозг | Промежуточный мозг | Конечный мозг | ||||
|
1. продолговатый мозг 2. мозжечок 3. Варолиев мост |
1. четверохолмие 2. ножки мозжечка |
1. зрительный бугор 2. подбугорье 3. надбугорье 4. забугорье |
1. большие полушария: кора, белое вещество, 2. подкорковые ядра |
||||
Рис 19А. Головной мозг
1. Лобная кора (когнитивная зона)
2. Двигательная кора
3. Зрительная кора
4. Мозжечок 5. Слуховая кора
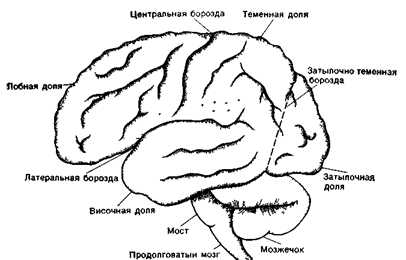
Рис 19Б. Вид сбоку
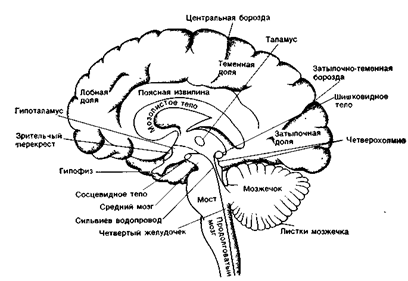
Рис 19В. Главные образования медальной поверхности головного мозга на средне-сагиттальном разрезе.
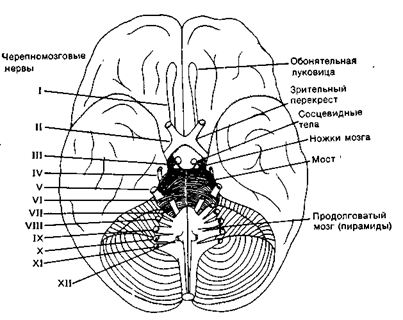
Рис 19Г. Нижняя поверхность головного мозга
Рис. 19. Строение головного мозга
Задний мозг
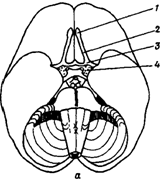
Задний мозг, включающий продолговатый мозг и Варолиев мост представляют собой филогенетически древнюю область центральной нервной системы, сохраняя черты сегментарного строения. В заднем мозге локализованы ядра и проводящие восходящие и нисходящие пути. По проводящим путям в задний мозг поступают афферентные волокна от вестибулярных и слуховых рецепторов, от рецепторов кожи и мышц головы, от рецепторов внутренних органов, а также от вышерасположенных структур головного мозга. В заднем мозге расположены ядра V-XII пар черепно-мозговых нервов, часть из которых иннервирует лицевую и глазодвигательную мускулатуру.
Продолговатый мозг
Продолговатый мозг расположен между спинным мозгом, Варолиевым мостом и мозжечком (рис. 20). На вентральной поверхности продолговатого мозга по средней линии проходит передняя срединная борозда, по ее бокам расположено два тяжа - пирамиды, сбоку от пирамид лежат оливы (рис. 20 А-В).
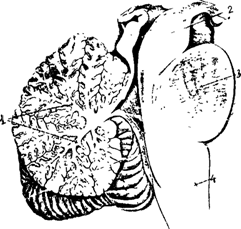
Рис. 20А. 1 - мозжечок 2- ножки мозжечка 3 - Мост 4 - Продолговатый мозг
|
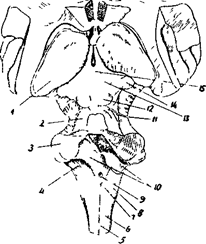
Рис 20Б. 1 - задняя часть таламуса 2,3,4- ножки мозжечка 5 - тонкий пучок Голя 6 - клиновидный пучок Бурдаха 7 - бугорок тонкого ядра 8 - бугорок клиновидного ядра 9 - отверстие ромбовидной ямки 10 - полость IV желудочка 11 - блоковидный нерв (IV) пара 12 – нижнее четверохолмие 13 - верхнее четверохолмие 14 - медиальное коленчатое тело 15 – эпфиз
|
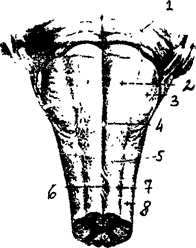
Рис. 20В. 1 - мост 2 - пирамида 3 - олива 4 - передняя серединная щель 5 - передняя боковая борозда 6 - пере крест переднего канатика 7 - передний канатик 8 -боковой канатик
Рис. 20. Продолговатый мозг
На задней стороне продолговатого мозга тянется задняя медиальная борозда. По ее бокам лежат задние канатики, которые идут к мозжечку в составе задних ножек.
Серое вещество продолговатого мозга
В продолговатом мозге расположены ядра четырех пар черепно-мозговых нервов. К ним относятся ядра языкоглоточного, блуждающего, добавочного и подъязычного нервов. Кроме того, выделяют нежное, клиновидное ядра и улитковые ядра слуховой системы, ядра нижних олив и ядра ретикулярной формации (гигантоклеточное, мелкоклеточное и латеральное), а также дыхательные ядра.
Ядра подъязычного (XII пара) и добавочного (XI пара) нервов - двигательные, иннервируют мышцы языка и мышцы, осуществляющие движение головы. Ядра блуждающего (X пара) и языкоглоточного (IX пара) нервов - смешанные, иннервируют мышцы глотки, гортани, щитовидную железу, осуществляют регуляцию глотания, жевания. Эти нервы состоят из афферентных волокон, идущих от рецепторов языка, гортани, трахеи и от рецепторов внутренних органов грудной и брюшной полости. Эфферентные нервные волокна иннервируют кишечник, сердце и сосуды.
Ядра ретикулярной формации не только активизируют кору больших полушарий, поддерживая сознание, но и образуют дыхательный центр, который обеспечивает дыхательные движения.
Таким образом, часть ядер продолговатого мозга регулирует жизненно важные функции (это ядра ретикулярной формации и ядра черепно-мозговых нервов). Другая часть ядер входит в состав восходящих и нисходящих путей (нежное и клиновидное ядра, улитковые ядра слуховой системы) ( рис. 21).
|
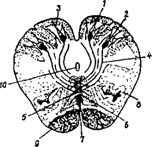
2 - клиновидное ядро;
3 - окончание волокон задних канатиков спинного мозга;
4 - внутренние дугообразные волокна - второй нейрон проприои пути коркового направления;
5 - перекрест петель находится в межоливном петлевом слое;
6 - медиальная петля - продолжение внутренних дугообразных вола
7 - шов, образован перекрестом петель;
8 - ядро оливы - промежуточное ядро равновесия;
9 - пирамидные пути;
10 - центральный канал.
Рис. 21. Внутреннее строение продолговатого мозга
Белое вещество продолговатого мозга
Белое вещество продолговатого мозга образовано длинными и короткими нервными волокнами
Длинные нервные волокна входят в состав нисходящих и восходящих проводящих путей. Короткие нервные волокна обеспечивают согласованную работу правой и левой половин продолговатого мозга.
Пирамиды продолговатого мозга - часть нисходящего пирамидного тракта, идущего в спинной мозг и оканчивающегося на вставочных нейронах и мотонейронах. Кроме того, через продолговатый мозг проходит рубро-спинальный путь. Нисходящие вестибуло-спинальный и ретикуло-спинальный тракты берут начало в продолговатом мозге соответственно от вестибулярных и ретикулярных ядер.
Восходящие спинно-мозжечковые тракты проходят через оливы продолговатого мозга и через ножки мозга и передают информацию от рецепторов опорно-двигательного аппарата к мозжечку.
Нежные и клиновидные ядра продолговатого мозга входят в состав одноименных путей спинного мозга, идущих через зрительные бугры промежуточного мозга до соматосенсорной коры.
Через улитковые слуховые ядра и через вестибулярные ядра проходят восходящие сенсорные пути от слуховых и вестибулярных рецепторов. В проекционную зону височной коры.
Таким образом, продолговатый мозг регулирует деятельность многих жизненно важных функций организма. Поэтому малейшие повреждение продолговатого мозга (травма, отек, кровоизлияние, опухоли), как правило, приводят к летальному исходу.
Варолиев мост
Мост представляет собой толстый валик, который граничит с продолговатым мозгом и ножками мозжечка. Восходящие и нисходящие пути продолговатого мозга проходят через мост, не прерываясь. В области соединения моста и продолговатого мозга выходит вестибулокохлеарный нерв (VIII пара). Вестибулокохлеарный нерв является чувствительным и передает информацию от слуховых и вестибулярных рецепторов внутреннего уха. Кроме того, в Варолиевом мосту расположены смешанные нервы, ядра тройничного нерва (V пара), отводящего нерва (VI пара), лицевого нерва (VII пара). Эти нервы иннервируют мышцы лица, кожу волосистой части головы, язык, боковые прямые мышцы глаза.
На поперечном срезе мост состоит из вентральной и дорсальной части - между ними граница - трапециевидное тело, волокна которого относят к слуховому пути. В области трапециевидного тела есть медиальное парабранхиальное ядро, которое связано с зубчатым ядром мозжечка. Собственное ядро моста осуществляется связь мозжечка с корой головного мозга. В дорсальной части моста лежат ядра ретикулярной формации и продолжаются восходящие и нисходящие пути продолговатого мозга.
Мост выполняет сложные и разнообразные функции, направленные на поддержание позы и сохранение равновесия тела в пространстве при изменении скорости движения.
Очень важным являются вестибулярные рефлексы, рефлекторные дуги которых проходят через мост. Они обеспечивают тонус шейных мышц, возбуждение вегетативных центров, дыхание, частоту сердечных сокращений, деятельность кишечно-сосудистого тракта.
Ядра тройничного, языкоглоточного, блуждающего нервов и моста связаны с захватом, пережевыванием и проглатыванием пищи.
Нейроны ретикулярной формации моста играют особую роль в активации коры больших полушарий и ограничении сенсорного притока нервных импульсов во время сна (рис. 22, 23)
|
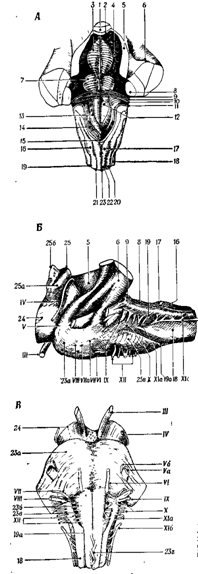
Рис. 22. Продолговатый мозг и мост.
А. Вид сверху (с дорсальной стороны).
Б. Вид сбоку.
В. Вид снизу(с вентральной стороны).
1 - язычок, 2 - передний мозговой парус, 3 - срединное возвышение, 4 - верхняя ямка, 5 - верхняя ножка мозжечка, 6 - средняя ножка мозжечка, 7 - лицевой бугорок, 8 - нижняя ножка мозжечка, 9 - слуховой бугорок, 10 - мозговые полосы, 11 - лента четвертого желудочка, 12 - треугольник подъязычного нерва, 13 - треугольник блуждающего нерва, 14 - area pos-terma, 15 - obex, 16 - бугорок клиновидного ядра, 17 - бугорок нежного ядра, 18 - боковой канатик, 19 - задняя боковая борозда, 19 а - передняя боковая борозда, 20 - клиновидный канатик, 21 - задняя промежуточная борозда, 22 - нежный канатик, 23 - задняя срединная борозда, 23 а - мост - основание), 23 б - пирамида продолговатого мозга, 23 в -олива, 23 г - перекрест пирамид, 24 - ножка мозга, 25 - нижний бугорок, 25 а - ручка нижнего бугорка, 256 - верхний бугорок
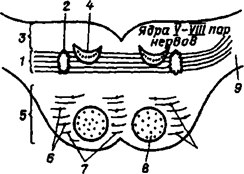
1 - трапецивидное тело 2 - ядро верхней оливы 3 - дорсальная содержит ядра VIII, VII, VI, V пар черепных нервов 4 – медальная часть моста 5 –-вентральная часть моста содержит собственные ядра и моста 7 - поперечные ядра моста 8 - пирамидные пути 9 – средняя ножка мозжечка.
Рис. 23. Схема внутреннего строения моста на вронтальном сечении
Мозжечок
Мозжечок представляет собой отдел мозга, расположенный позади полушарий большого мозга над продолговатым мозгом и мостом.
Анатомически в мозжечке выделяют среднюю часть - червь, и два полушария. С помощью трех пар ножек (нижних, средних и верхних) мозжечок связан со стволом мозга. Нижние ножки соединяют мозжечок с продолговатым и спинным мозгом, средние - с мостом, а верхние со средним и промежуточным мозгом ( рис. 24 ).
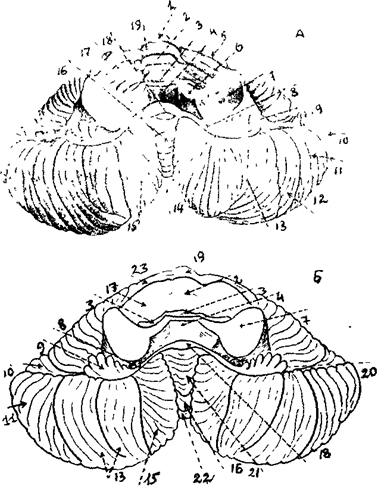
1 - червь 2 - центральная долька 3 - язычок червя 4 - передний парус мозжечка 5 - верхнее полушарие 6 - передняя ножка мозжечка 8 - ножка клочка 9 – клочок 10 - верхняя полулунная долька 11 - нижняя полулунная долька 12 - нижнее полушарие 13 - двубрюшная долька 14 - долька мозжечка 15 - миндалина мозжечка 16 - пирамида червя 17 - крыло центральной дольки 18 - узелок 19 - верхушка 20 - бороздка 21 - втулочка червя 22 - бугор червя 23 - четырехугольная долька.
Рис. 24. Внутреннее строение мозжечка
Мозжечок построен по ядерному типу - поверхность полушарий представлена серым веществом, составляющим новую кору. Кора образует извилины, которые отделяются друг от друга бороздами. Под корой мозжечка расположено белое вещество, в толще которого выделяют парные ядра мозжечка (рис. 25). К ним относят ядра шатра, шаровидное ядро, пробковое ядро, зубчатое ядро. Ядра шатра связаны с вестибулярным аппаратом, шаровидное и пробковое ядра с движение туловища, зубчатое ядро – с движением конечностей.
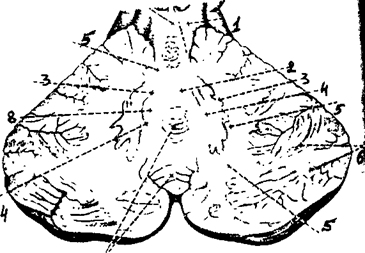
1- передние ножки мозжечка; 2 - ядра шатра; 3 - зубчатое ядро; 4 - пробковидное ядро; 5 - белая субстанция; 6 - полушария мозжечка; 7 – червь; 8 шаровидное ядро
Рис. 25. Ядра мозжечка
Кора мозжечка однотипна и состоит из трех слоев: молекулярного, ганглиозного и гранулярного, в которых находятся 5 типов клеток: клетки Пуркинье, корзинчатые, звездчатые, гранулярные и клетки Гольджи (рис.26). В поверхностном, молекулярном слое, расположены дендритные разветвления клеток Пуркинье, представляющие собой один из наиболее сложно устроенных нейронов мозга. Дендритные отростки обильно покрыты шипиками, что свидетельствует о большом количестве синапсов. Кроме клеток Пуркинье в этом слое много аксонов параллельных нервных волокон (Т-образно ветвящиеся аксоны гранулярных клеток). В нижней части молекулярного слоя находятся тела корзинчатых клеток, аксоны которых образуют синаптические контакты в области аксонных холмиков клеток Пуркинье. В молекулярном слое имеются еще и звездчатые клетки.

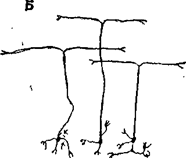
А. Клетка Пуркинье. Б. Клетки-зерна.
 В. Клетка Гольджи.
В. Клетка Гольджи.
Рис. 26. Типы нейронов мозжечка.
Под молекулярным слоем расположен ганглиозный слой, в котором находятся тела клеток Пуркинье.
Третий слой – гранулярный – представлен телами вставочных нейронов (клеток-зерен или гранулярных клеток). В гранулярном слое имеются еще клетки Гольджи, аксоны которых поднимаются в молекулярный слой.
В кору мозжечка поступают только два типа афферентных волокон: лазающие и мшистые, по которым в мозжечок приходят нервные импульсы. Каждое лазающее волокно имеет контакт с одной клеткой Пуркинье. Разветвления мшистого волокна образуют контакты в основном с гранулярными нейронами, но не контактируют с клетками Пуркинье. Синапсы мшистого волокна являются возбуждающими (рис. 27).
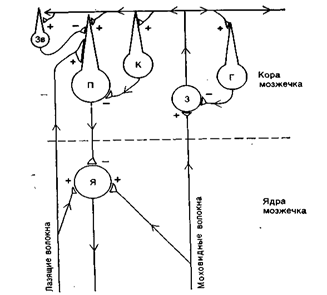
К коре и ядрам мозжечка поступают возбуждающие импульсы как по лазящим, так и моховидным волокнам. Из мозжечка же сигналы идут лишь от клеток Пуркинье (П), угнетающих активность нейронов в ядрах1 мозжечка (Я). К собственным нейронам коры мозжечка относятся возбуждащщие клетки-зерна (3) и тормозные корзинчатые нейроны (К), нейроны Гольджи (Г) и звездчатые нейроны (Зв). Стрелками указано направление движения нервных импульсов. Имеются как возбуждающие (+), так и ;тормозные (-) синапсы.
Рис. 27. Нервная цепь мозжечка.
Таким образом, в кору мозжечка входят два типа афферентных волокон: лазающие и мшистые. По этим волокнам передается информация от тактильных рецепторов и рецепторов опорно-двигательного аппарата, а также от всех структур мозга, регулирующих двигательную функцию организма.
Эфферентное влияние мозжечка осуществляется через аксоны клеток Пуркинье, которые являются тормозными. Аксоны клеток Пуркинье оказывают свое влияние действие либо непосредственно на мотонейроны спинного мозга, либо косвенно через нейроны ядер мозжечка или другие двигательные центры.
У человека в связи с прямохождением и трудовой деятельностью мозжечок и его полушария достигают наибольшего развития и размера.
При повреждении мозжечка наблюдаются нарушения равновесия и мышечного тонуса. Характер нарушений зависит от места повреждений. Так, при поражении ядер шатра нарушается равновесие тела. Это проявляется в шатающейся походке. При повреждении червя, пробкового и шаровидного ядер – нарушается работа мышц шеи и туловища. У больного возникают трудности при приеме пищи. При поражении полушарий и зубчатого ядра – работа мышц конечностей (тремор), затрудняется его профессиональная деятельность.
Кроме того, у всех больных с повреждением мозжечка в связи с нарушением координации движений и тремором (дрожание) быстро возникает утомление.
Средний мозг
Средний мозг, как и продолговатый и Варолиев мост, относится к стволовым структурам ( рис. 28).
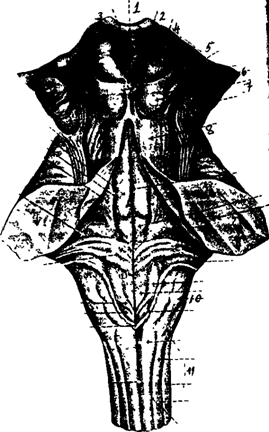
1 - комисура поводков
2 - поводок
3 - шишковидная железа
4 - верхнее двухолмие среднего мозга
5 - медиальное коленчатое тело
6 - латеральное коленчатое тело
7 - нижнее двухолмие среднего мозга
8 - верхние ножки мозжечка
9 - средние ножки мозжечка
10 - нижние ножки мозжечка
11- продолговатый мозг
Рис. 28. Задний мозг
Средний мозг состоит из двух частей: крыши мозга и ножек мозга. Крыша среднего мозга представлена четверохолмием, в котором выделяют верхние и нижние бугры. В толще ножек мозга выделяют парные скопления ядер, получивших названия черная субстанция и красное ядро. Через средний мозг проходят восходящие пути к промежуточному мозгу и мозжечку и нисходящие пути - из коры больших полушарий, подкорковых ядер и промежуточного мозга до ядер продолговатого и спинного мозга.
В нижних буграх четверохолмия располагаются нейроны, получающие афферентные сигналы от слуховых рецепторов. Поэтому нижние бугры четверохолмия называют первичным слуховым центром. Через первичный слуховой центр проходит рефлекторная дуга ориентировочного слухового рефлекса, который проявляется в повороте головы в сторону акустического сигнала.
Верхние бугры четверохолмия являются первичным зрительным центром. На нейроны первичного зрительного центра поступают афферентные импульсы от фоторецепторов. Верхние бугры четверохолмия обеспечивают ориентировочный зрительный рефлекс – поворот головы в сторону зрительного стимула.
В осуществлении ориентировочных рефлексов принимают участие ядра бокового и глазодвигательного нервов, которые иннервируют мышцы глазного яблока, обеспечивая его движение.
Красное ядро содержит нейроны разных размеров. От крупных нейронов красного ядра начинается нисходящий рубро-спинальный тракт, который оказывает действие на мотонейроны, тонко регулирует мышечный тонус.
Нейроны черной субстанции содержат пигмент меланин и придают этому ядру темный цвет. Черная субстанция, в свою очередь, посылает сигналы к нейронам ретикулярных ядер ствола мозга и подкорковым ядрам.
Черная субстанция участвует в сложной координации движений. В ней содержатся дофаминергические нейроны, т.е. выделяющие в качестве медиатора – дофамин. Одна часть таких нейронов регулирует эмоциональное поведение, другая – играет важную роль в контроле сложных двигательных актов. Повреждение черной субстанции, приводящее к дегенерации дофаминергических волокон обуславливает неспособность приступить к выполнению произвольных движений головы и рук, когда больной сидит спокойно (болезнь Паркинсона) ( рис. 29 А,Б ).
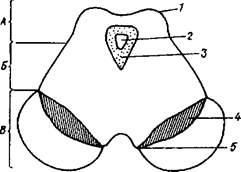
Рис. 29А. 1 - холмик 2 - водопровод большого мозга 3 - центральное серое вещество 4 - черная субстанция 5 - медиальная борозда ножки большого мозга
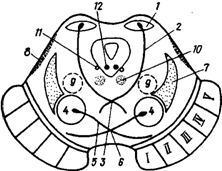
Рис. 29Б. Схема внутреннего строения среднего мозга на уровне нижних холмиков (фронтальное сечение)
1 - ядро нижнего холмика, 2 - двигательный путь экстрапирамидной системы, 3 - дорсальный перекрест покрышки, 4 - красное ядро, 5 - красноядерный - спинномозговой путь, 6 - вентральный перекрест покрышки, 7 - медиальная петля, 8 - латеральная петля, 9 - ретикулярная формация, 10 - медиальный продольный пучок, 11 - ядро среднемозгового тракта тройничного нерва, 12 - ядро бокового нерва, I-V - нисходящие двигательные пути ножки мозга
Рис. 29. Схема внутреннего строения среднего мозга
Промежуточный мозг
Промежуточный мозг образует стенки III желудочка. Главными структурами его являются зрительные бугры (таламус) и подбугровая область (гипоталамус), а также надбугровая область (эпиталамус) (рис. 30 А,Б).
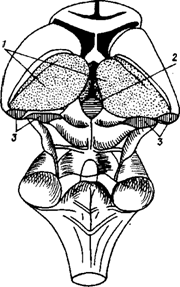
Рис. 30 А. 1 - thalamus (зрительный бугор) - подкорковый центр всех видов чувствительности, "чувствилище" мозга; 2 - epithalamus (надбугорная область); 3 - metathalamus (забугорная область).
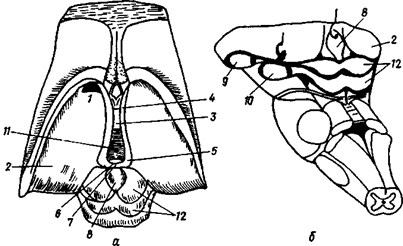
Рис. 30 Б. Схемы зрительного мозга (thalamencephalon): а - вид сверху б - вид сзади и снизу.
Таламус (зрительный бугор) 1 - передний бурф зрительного бугра, 2 - подушка 3 - межбугорное сращение 4 - мозговая полоска зрительного бугра
Эпиталамус (надбугровая область) 5 - треугольник поводка, 6 - поводок, 7 - спайка поводка, 8 - шишковидное тело (эпифиз)
Метаталамус (забугорная область) 9 - латеральное коленчатое тело, 10 - медиальное коленчатое тело, 11 - III желудочек, 12 - крыша среднего мозга
Рис. 30. Зрительный мозг
В глубине мозговой ткани промежуточного мозга расположены ядра наружных и внутренних коленчатых тел. Наружная граница образована белым веществом, отделяющим промежуточный мозг от конечного.
Таламус (зрительные бугры)
Нейроны таламуса образуют 40 ядер. Топографически ядра таламуса подразделяются на передние, срединные и задние. Функционально эти ядра можно разделить на две группы: специфические и неспецифические.
Специфические ядра входят в состав специфических проводящих путей. Это восходящие пути, которые передают информацию от рецепторов органов чувств к проекционным зонам коры полушарий большого мозга.
Важнейшими из специфических ядер являются латеральное коленчатое тело, участвующее в передаче сигналов от фоторецепторов и медиальное коленчатое тело, передающее сигналы от слуховых рецепторов.
Неспецифические яра таламуса относят к ретикулярной формации. Они выполняют роль интегративных центров и оказывают преимущественно активирующее восходящее влияние на кору полушарий большого мозга( рис. 31 А,Б )
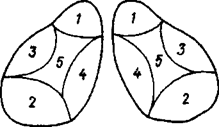
Рис. 31 А.
1 - передняя группа (обонятельные); 2 - задняя группа (зрительные); 3 - латеральная группа (общая чувствительность); 4 - медиальная группа (экстрапирамидная система; 5 - центральная группа (ретикулярная формация).
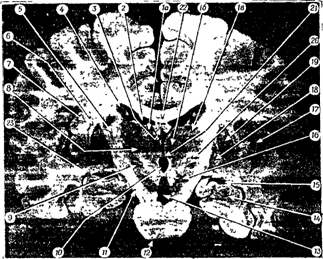
Рис. 31Б. Фронтальный срез головного мозга на уровне середины зрительного бугра. 1а - переднее ядро зрительного бугра. 16 - медиальное ядро зрительного бугра, 1в - латеральное ядро зрительного бугра, 2 - боковой желудочек, 3 - свод, 4 - хвостатое ядро, 5 - внутренняя капсула, 6 - наружная капсула, 7 - внешняя капсула (capsula extrema), 8 - вентральное ядро зрительного бугра, 9 - субталамическое ядро, 10 - третий желудочек, 11 - ножка мозга. 12 - мост, 13 - межножковая ямка, 14 - ножка гиппокампа, 15 - нижний рог бокового желудочка. 16 - черное вещество, 17 - островок. 18 - бледный шар, 19 - скорлупа, 20 - поля Фореля Н; и ЬЬ. 21 - межталамическое сращение, 22 - мозолистое тело, 23 - хвост хвостатого ядра.
Рис 31. Схема групп ядер зрительного бугра
Активацию нейронов неспецифических ядер таламуса особенно эффективно вызывают болевые сигналы (таламус – высший центр болевой чувствительности).
Повреждения неспецифических ядер таламуса приводят также к нарушению сознания: потерей активной связи организма с окружающей средой.
Подбугорье (гипоталамус)
Гипоталамус образован группой ядер, ррасположенных у основания мозга. Ядра гипоталамуса представляют собой подкорковые центры автономной нервной системы всех жизненно важных функций организма.
Топографически гипоталамус разделяется на преоптическую область, области переднего, среднего и заднего гипоталамуса. Все ядра гипоталамуса парные ( рисю 32 А-Г).
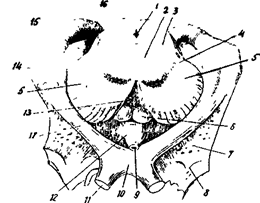
1 - водопровод 2 - красное ядро 3 - покрышка 4 - черная субстанция 5 - ножка мозга 6 -сосцевидные тела 7 - переднее продырявленное вещество 8 - обобнятельный треугольник 9 - воронка 10 - зрительный перекрест 11. зрительный нерв 12 - серый бугор 13-заднее продырявленное вещество 14 - наружное коленчатое тело 15 - медиальное коленчатое тело 16 - подушка 17 - зрительный тракт
Рис. 32А. Метаталамус и гипоталамус
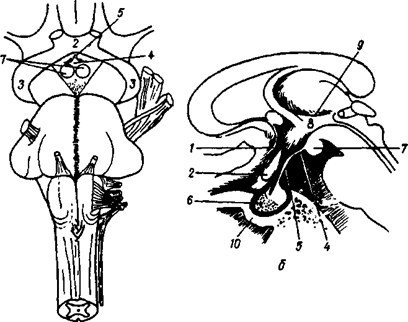
а - вид снизу; б - серединное сагиттальное сечение.
Зрительная часть (pars optica): 1 - конечная пластинка; 2 - зрительный перекрест; 3 - зрительный тракт; 4 - серый бугор; 5 - воронка; 6 - гипофиз;
Обонятельная часть: 7 - сосковидные тела - подкорковые обонятельные центры; 8 - подбугорная область в узком смысле слова является продолжением ножек мозга, содержит черное вещество, красное ядро и Люисово тело, которое является звеном экстрапирамидной системы и вегетативным центром; 9 - подбугорная Монроева борозда; 10 - турецкое седло, в ямке которого находится гипофиз.
Рис. 32Б. Подбугорная область (hypothalamus)
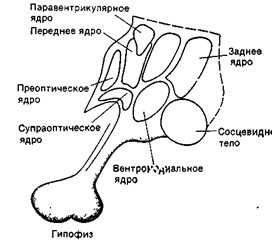
Рис. 32В. Главные ядра гипоталамуса
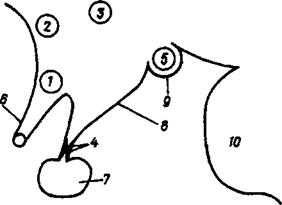
1 - nucleus supraopticus; 2 - nucleus preopticus; 3 - nuclius paraventricularis; 4 - nucleus infundibularus; 5 - nucleus corporis mamillaris; 6 - зрительный перекрест; 7 - гипофиз; 8 - серый бугор; 9 - сосцевидное тело; 10 мост.
Рис. 32Г. Схема нейросекреторных ядер подбугорной обласп (Hypothalamus)
В преоптическую область входят перивентрикулярное, медиальное и латеральное преоптические ядра.
В группу переднего гипоталамуса относят супраоптическое, супрахиазматическое и паравентрикулярное ядра.
Средний гипоталамус составляет вентромедиальное и дорсомедиальное ядра.
В заднем гипоталамусе различают заднее гипоталамическое, перифорникальное и мамиллярное ядра.
Связи гипоталамуса обширны и сложны. Афферентные сигналы в гипоталамус поступают от коры больших полушарий, подкорковых ядер и от таламуса. Основные эфферентные пути доходят до среднего мозга, таламуса и подкорковых ядер.
Гипоталамус является высшим центром регуляции сердечно-сосудистой системы, водно-солевого, белкового, жирового, углеводного обменов. В этой области мозга расположены центры, связанные с регуляцией пищевого поведения. Важная роль гипоталамуса – регуляция. Электрическое раздражение задних ядер гипоталамуса приводит к гипертермии, в результате повышения обмена веществ.
Гипоталамус принимает также участие в поддержании биоритма “сон-бодрствование”.
Ядра переднего гипоталамуса связаны с гипофизом и осуществляют транспорт биологически активных веществ, которые вырабатываются нейронами этих ядер. Нейроны преоптического ядра вырабатывают рилизинг-факторы (статины и либерины), контролирующие синтез и высвобождение гормонов гипофиза.
Нейроны преоптического, супраоптического, паравентрикулярного ядер вырабатывают истинные гормоны – вазопрессин и окситоцин, которые по аксонам нейронов спускаются в нейрогипофиз, где хранятся до высвобождения – поступления в кровь.
Нейроны передней доли гипофиза вырабатывают 4 вида гормонов: 1) соматотропный гормон, регулирующий рост; 2) гонадотропный гормон, способствующий росту половых клеток, желтого тела, усиливает выработку молока; 3) тиреотропный гормон – стимулирует функцию щитовидной железы; 4) адренокортикотропный гормон – усиливает синтез гормонов коры надпочечников.
Промежуточная доля гипофиза выделяет гормон интермедин, влияющий на пигментацию кожи.
Задняя доля гипофиза выделяет два гормона – вазопрессин, влияющий на гладкую мускулатуру артериол, и окситоцин – действует на гладкую мускулатуру матки и стимулирует выделение молока.
Гипоталамус играет также важную роль в эмоциональном и половом поведении.
В состав эпиталамуса (шишковидная железа) входит эпифиз. Гормон эпифиза – мелатонин – тормозит в гипофизе образование гонадотропных гормонов, а это в свою очередь задерживает половое развитие.
Передний мозг
Передний мозг состоит из трех анатомически обособленных частей – коры полушарий большого мозга, белого вещества и подкорковых ядер.
В соответствии с филогенезом коры полушарий большого мозга выделяют древнюю кору (архикортекс), старую кору (палеокортекс) и новую кору (неокортекс). К древней коре относят обонятельные луковицы, в которые поступают афферентные волокна от обонятельного эпителия, обонятельные тракты – расположенные на нижней поверхности лобной доли и обонятельные бугорки – вторичные обонятельные центры.
Старая кора включает кору поясной извилины, кору гиппокампа и миндалину.
Все остальные области коры являются новой корой. Древнюю и старую кору называют обонятельным мозгом (рис. 33).
Обонятельный мозг, помимо функций, связанных с обонянием, обеспечивает реакции настораживания и внимания, принимает участие в регуляции вегетативных функций организма. Эта система играет также важную роль в осуществлении инстинктивных форм поведения (пищевого, полового, оборонительного) и формировании эмоций.
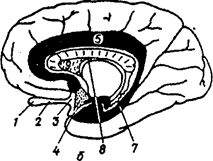
а - вид снизу; б - на сагиттальном сечении мозга
Переферический отдел: 1 - bulbus olfactorius (обонятельная луковица; 2 - tractus olfactories (обонятельный путь); 3 - trigonum olfactorium (обонятельный треугольник); 4 - substantia perforate anterior (переднее продырявленное вещество).
Центральный отдел - извилины мозга: 5 - сводчатая извилина; 6 - hippocampus расположен в полости нижнего рога бокового желудочка; 7 - продолжение серого облачения мозолистого тела; 8 - свод; 9 - прозрачная перегородка проводящие пути обонятельного мозга.
Рис 33. Обонятельный мозг
Раздражение структур старой коры оказывает влияние на сердечно-сосудистую систему и дыхание, вызывает гиперсексуальность, изменяет эмоциональное поведение.
При электрическом раздражении миндалины наблюдаются эффекты, связанные с деятельностью пищеварительного тракта: облизывание, жевание, глотание, изменение перистальтики кишечника. Раздражение миндалины влияет и на деятельность внутренних органов – почек, мочевого пузыря, матки.
Таким образом, существует связь структур старой коры с вегетативной нервной системой, с процессам, направленными на поддержание гомеостаза внутренних сред организма.
Конечный мозг
В состав конечного мозга входят: кора полушарий, белое вещество и расположенные в его толще подкорковые ядра.
Поверхность полушарий большого мозга складчатая. Борозды – углубления делят ее на доли.
Центральная (Роландова) борозда отделяет лобную долю от теменной доли. Боковая (Сильвиева) борозда отделяет височную долю от теменной и лобной долей. Затылочно-теменная борозда образует границу между теменной, затылочной и височной долями ( рис.34 А,Б, рис. 35)
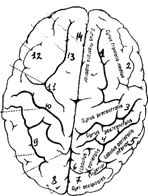

1 - верхняя лобная извилина; 2 - средняя лобная извилина; 3 - предцентральная извилина; 4 - постцентральная извилина; 5 - нижняя теменная извилина; 6 - верхняя теменная извилина; 7 - затылочная извилина; 8 - затылочная бороздка; 9 - внутритеменная борозда; 10 - центральная борозда; 11 - предцентральная извилина; 12 - нижняя лобная борозда; 13 - верхняя лобная борозда; 14 - вертикальная щель.
Рис. 34А. Головной мозг с дорсальной поверхности
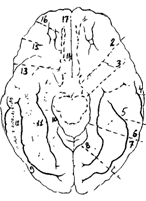
1 - обонятельная борозда; 2 - передняя продырявленная субстанция; 3 - крючок; 4 - средняя височная борозда; 5 - нижняя височная борозда; 6 - борозда морского конька; 7 - окольная борозда; 8 - шпорная борозда; 9 - клин; 10 - парагиппокампальная извилина; 11 - затылочно-височная борозда; 12 - нижне-теменная извилина; 13 - обонятельный треугольник; 14 - прямая извилина; 15 - обонятельный тракт; 16 - обонятельная луковица; 17 - вертикальная щель.
Рис. 34Б. Головной мозг с вентральной поверхности
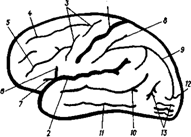
1 - центральная борозда (Роланда); 2 - латеральная борозда (Сильвиева борозда); 3 - предцентральная борозда; 4 - верхняя лобная борозда; 5 - нижняя лобная борозда; 6 - восходящая ветвь; 7 - передняя ветвь; 8 - зацентральная борозда; 9 - внутритеменная борозда; 10- верхняя височная борозда; 11 - нижняя височная борозда; 12 - поперечная затылочная борозда; 13 - затылочная борозда.
Рис. 35. Борозды верхнелатеральной поверхности полушария (левая сторона)
Таким образом, борозды разделяют полушария конечного мозга на пять долей: лобную, теменную, височную, затылочную и островковую долю, которая расположена под височной долей (рис. 36).
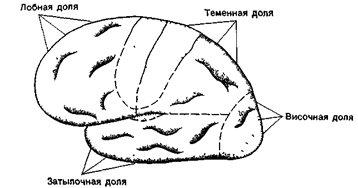
Рис. 36. Проекционные (отмечены точками) и ассоциативные (светлые) зоны коры головного мозга. К проекционным зонам относятся двигательная область (лобная доля), соматосенсорная область (теменная доля), зрительная область (затылочная доля) и слуховая область (височная доля).
На поверхности каждой доли также расположены борозды.
Различают борозды трех порядков: первичные, вторичные и третичные. Первичные борозды относительно стабильные и наиболее глубокие. Это границы крупных морфологических отделов мозга. Вторичные борозды отходят от первичных, а третичные от вторичных.
Между бороздами имеются складки – извилины, форма которых определяется конфигурацией борозд.
В лобной доле выделяют верхнюю, среднюю и нижнюю лобные извилины. В височной доле имеются верхняя, средняя и нижняя височные извилин. Передняя центральная извилина (прецентральная) расположена перед центральной бороздой. Задняя центральная извилина (постцентральная) находится за центральной бороздой.
У человека наблюдается большая вариабельность борозд и извилин конечного мозга. Несмотря на эту индивидуальную изменчивость внешнего строения полушарий, это не сказывается на структуре личности и сознания.
Цитоархитектоника и миелоархитектоника новой коры
В соответствии с делением полушарий на пять долей выделяют пять основных областей – лобную, теменную, височную, затылочную и островковую, имеющие различия в строении и выполняющие разные функции. Однако общий план строения новой коры одинаков. Новая кора – это слоистая структура (рис. 37). I - молекулярный слой, образован преимущественно нервными волокнами, идущими параллельно поверхности. Среди параллельных волокон расположено небольшое количество зернистых клеток. Под молекулярным слоем расположен II слой – наружный зернистый. III слой – наружный пирамидный, IV слой, внутренний зернистый, V слой – внутренний пирамидный и VI слой – мультиформный. Название слоев дано по названию нейронов. Соответственно, во II и IV слоях – сомы нейронов имеют округлую форму (клетки-зерна) (наружный и внутренний зернистый слои), а в III и IV слоях сомы имеют пирамидную форму (в наружном пирамидном – малые пирамиды, а во внутреннем пирамидном – большие пирамиды или клетки Беца). VI слой характеризуется наличием нейронов разнообразной формы (веретенообразной, треугольной и др.).
Главные афферентные входы в кору полушарий большого мозга – нервные волокна, идущие из таламуса. Корковые нейроны, которые воспринимают афферентные импульсы, идущие по этим волокнам, называют сенсорными, а зона, где расположены сенсорные нейроны – проекционными зонами коры.
Главные эфферентные выходы из коры – аксоны пирамид V слоя. Это эфферентные, двигательные нейроны, участвующие в регуляции двигательных функций. Большинство нейронов коры – вставочные, участвующие в переработке информации и обеспечивающие межкортикальные связи.

Типичные нейроны коры
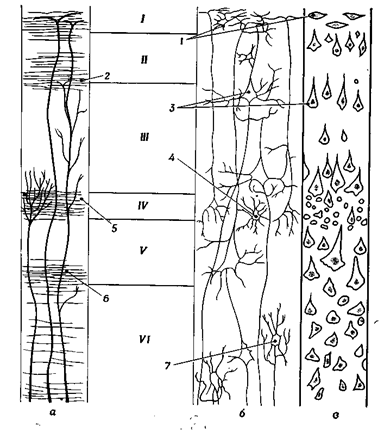
Римскими цифрами обозначены клеточные слои I - молекулярный сдой; II - наружный зернистый слой; III - наружный пирамидный слой; IV - внутренний зернистый слой; V - внутренний приамидный слой; VI-мультиформный слой.
а - афферентные волокна; б - типы клеток, выявляемые на препаратах, импрегнированных по методу Голдбжи; в - цитоархитектоника, выявляемая при окрашивании по Нисслю. 1 - горизонтальные клетки, 2 - полоска Кеса, 3 - пирамидные клетки, 4 – звездчатые клетки, 5 - наружная полоска Белларже, 6 - внутренняя полоска Белларже, 7 - видоизмененная пирамидная клетка.
Рис. 37. Цитоархитектоника (А) и миелоархитектоника (Б) коры полушарий большого мозга.
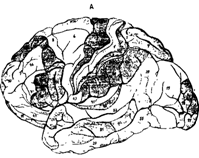
При сохранении общего плана строения было установлено, что разные участки коры (в пределах одной области) отличаются по толщине слоев. В некоторых слоях можно выделить несколько подслоев. Кроме того, имеются различия клеточного состава (разнообразие нейронов, плотность и их расположение). С учетом всех этих различий Бродман выделил 52 участка, которые назвал цитоархитектоническими полями и обозначил арабскими цифрами от 1 до 52 (рис.38 А,Б ).
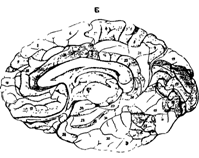
А вид сбоку. Б срединно-сагиттальный; срез.
Рис. 38. Схема расположения полей по Бордману
Каждое цитоархитектоническое поле отличается не только клеточным строением, но и расположением нервных волокон, которые могут идти как в вертикальном, так и в горизонтальном направлениях. Скопление нервных волокон в пределах цитоархитектонического поля называют миелоархитектоникой.
В настоящее время все большее признание находит “колончатый принцип” организации проекционных зон коры.
Согласно этому принципу каждая проекционная зона состоит из большого количества вертикально ориентированных колонок, диаметром приблизительно в 1 мм. Каждая колонка объединяет около 100 нейронов, среди которых имеются сенсорные, вставочные и эфферентные нейроны, связанные между собой синаптическими связями. Одиночная “корковая колонка” участвует в переработке информации от ограниченного количества рецепторов, т.е. выполняет специфическую функцию.
Система волокон полушарий
Оба полушария имеют три типа волокон. По проекционным волокнам возбуждение поступает в кору от рецепторов по специфическим проводящим путям. Ассоциативные волокна связывают между собой различные области одного и того же полушария. Например, затылочную область с височной, затылочную – с лобной, лобную – с теменной областью. Комиссуральные волокна связывают симметричные участки обоих полушарий. Среди комиссуральных волокон выделяют: переднюю, заднюю мозговые спайки и мозолистое тело ( рис. 39 А.Б).
|
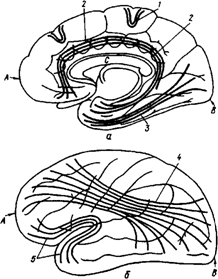
Рис. 39А. а - медиальная поверхность полушария;
б - верхнеальтеральная поверхность полушария;
А - лобный полюс;
В - затылочный полюс;
С - мозолистое тело;
1 - дугообразные волокна большого мозга соединяют между собой соседние извилины;
2 - пояс - пучок обонятельного мозга лежит под сводчатой извилиной, простирается из области обонятельного треугольника до крючка;
3 - нижний продольный пучок связывает затылочную и височную область;
4 - верхний продольный пучок связывает лобную, затылочную, височную доли и нижнетеменную дольку;
5 - крючковидный пучек расположен у переднего края островка и соединяет лобный полюс с височным.
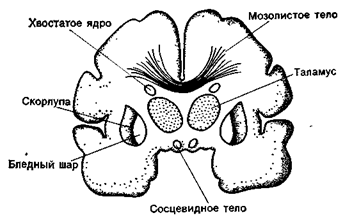
Рис. 39Б. Кора головного мозга на поперечном разрезе. Оба полушария соединены пучками белого вещества, образующими мозолистое тело (комиссуральные волокна).
Рис. 39. Схема ассоциативных волокон
Ретикулярная формация
Ретикулярная формация (сетчатое вещество мозга) была описана анатомами в конце прошлого века.
Ретикулярная формация начинается в спинном мозге, где она представлена желатинозной субстанцией основания заднего мозга. Основная ее часть находится в центральном стволе мозга и в промежуточном мозге. Она состоит из нейронов различной формы и размеров, которые имеют обширные ветвящиеся отростки, идущие в разных направлениях. Среди отростков выделяют короткие и длинные нервные волокна. Короткие отростки обеспечивают локальные связи, длинные - формируют восходящие и нисходящие пути ретикулярной формации.
Скопления нейронов образуют ядра, которые находятся на разных уровнях мозга (спинном, продолговатом, среднем, промежуточном). Большинство ядре ретикулярной формации не имеют четких морфологических границ и нейроны этих ядер объединяются только по функциональному признаку (дыхательный, сердечно-сосудистый центр и др.). Однако на уровне продолговатого мозга выделяют ядра с четко обозначенными границами - ретикулярное гигантоклеточное, ретикулярное мелкоклеточное и латеральное ядра. Ядра ретикулярной формации моста по существу являются продолжением ядер ретикулярной формации продолговатого мозга. Наиболее крупные из них - каудальное, медиальное и оральное ядра. Последнее переходит в клеточную группу ядер ретикулярной формации среднего мозга и ретикулярное ядро покрышки мозга. Клетки ретикулярной формации являются началом как восходящих, так и нисходящих путей, дающих многочисленные коллатерали (окончания), которые образуют синапсы на нейронах разных ядер центральной нервной системы.
Волокна ретикулярных клеток, направляющихся в спинной мозг, образуют ретикулоспинальный тракт. Волокна восходящих трактов, начинающихся в спинном мозге, связывают ретикулярную формацию с мозжечком, средним мозгом, промежуточным мозгом и корой полушарий большого мозга.
Выделяют специфические и неспецифические ретикулярной формации. Например, некоторая часть восходящих путей ретикулярной формации получают коллатерали от специфических путей (зрительных, слуховых и т.д.), по которым афферентные импульсы передаются в проекционные зоны коры.
Неспецифические восходящие и нисходящие пути ретикулярной формации влияют на возбудимость различных отделов мозга, в первую очередь коры полушарий большого мозга и спинной мозг. Эти влияния по функциональному значению могут быть как активирующими, так и тормозными, поэтому выделяют: 1) восходящее активирующее влияние, 2) восходящее тормозное влияние, 3) нисходящее активирующее влияние, 4) нисходящее тормозное влияние. На основании этих факторов ретикулярную формацию рассматривают как регулирующую неспецифическую систему мозга.
Наиболее изучено активирующее влияние ретикулярной формации на кору полушарий большого мозга. Большинство восходящих волокон ретикулярной формации диффузно оканчивается в коре полушарий и поддерживает ее тонус, обеспечивает внимание. Примером тормозных нисходящих влияний ретикулярной формации является снижение тонуса скелетных мышц человека во время определенных стадий сна.
Нейроны ретикулярной формации чрезвычайно чувствительны к гуморальным веществам. Это опосредованный механизм влияния различных гуморальных факторов и эндокринной системы на высшие отделы мозга. Следовательно, тонические воздействия ретикулярной формации зависят от состояния всего организма (рис.40).
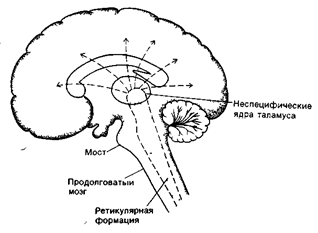
Рис. 40. Активирующая ретикулярная система (АРС) — нервная сеть, по которой сенсорное возбуждение передается от ретикулярной формации ствола мозга к неспецифическим ядрам таламуса. Волокна от этих ядер регулируют уровень активности коры.
Подкорковые ядра
Подкорковые ядра входят в состав конечного мозга и расположены внутри белого вещества полушарий большого мозга. К ним относят хвостатое тело и скорлупу, объединяемые под общим названием “полосатое тело” (стриатум) и бледный шар, состоящий из чечевицеобразного тела, шелухи и миндалины. Подкорковые ядра и ядра среднего мозга (красное ядро и черная субстанция) составляют систему базальных ганглиев (ядер)(рис.41). К базальным ганглиям поступают импульсы от двигательной коры и мозжечка. В свою очередь, сигналы от базальных ганглиев направляются к двигательной коре, мозжечку и ретикулярной формации, т.е. существуют две нейронные петли: одна связывает базальные ганглии с двигательной корой, другая - с мозжечком.
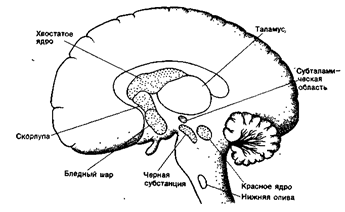
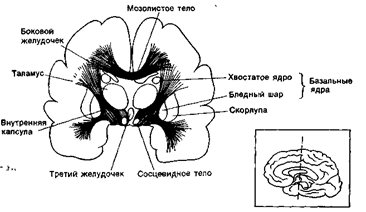
Рис. 41. Система базальных ганглиев
Подкорковые ядра принимают участие в регуляции двигательной активности, регулируя сложные движения при ходьбе, поддержании позы, при еде. Они организуют медленные движения (перешагивание через препятствия, вдевание нитки в иголку и т.д.).
Имеются данные, что полосатое тело участвует в процессах запоминания двигательных программ, так как раздражение этой структуры приводит к нарушению обучения и памяти. Полосатое тело оказывает тормозное влияние на различные проявления двигательной активности и на эмоциональные компоненты двигательного поведения, в частности на агрессивные реакции.
Основными медиаторами базальных ганглиев являются: дофамин (особенно в черной субстанции) и ацетилхолин. Поражение базальных ганглиев вызывает медленные извивающиеся непроизвольные движения, на фоне которых возникают резкие сокращения мышц. Непроизвольные порывистые движения головы и конечностей. Болезнь Паркинсона, основными симптомами которой является тремор (дрожание) и мышечная ригидность (резкое повышение тонуса мышц-разгибателей). Из-за ригидности больной с трудом может начать движение. Постоянный тремор препятствует выполнению мелких движений. Болезнь Паркинсона возникает при поражении черной субстанции. В норме черная субстанция оказывает тормозное влияние на хвостатое ядро, скорлупу и бледный шар. При ее разрушении тормозные влияния устраняются, в результате чего усиливается возбуждающее базальных ганглиев на кору головного мозга и ретикулярную формацию, что вызывает характерные симптомы болезни.
Лимбическая система
Лимбическая система представлена расположенными на границе отделами новой коры (неокортекса) и промежуточного мозга. Она объединяет комплексы структур разного филогенетического возраста, часть из которых является корковыми, а часть - ядерными.
К корковым структурами лимбической системы относят гиппокампальную, парагиппокампальную и поясную извилины (старя кора). Древняя кора представлена обонятельной луковицей и обонятельными бугорками. Новая кора - часть лобной, островковой и височной коры.
Ядерные структуры лимбической системы объединяют миндалину и септальные ядра и передние таламические ядра. Многие анатомы причисляют к лимбической системе преоптическую область гипоталамуса и маммилярные тела. Структуры лимбической системы образуют 2-х сторонние связи и связаны с другими отделами головного мозга.
Лимбическая система контролирует эмоциональное поведение и регулирует эндогенные факторы, обеспечивающие мотивации. Положительные эмоции связаны преимущественно с возбуждением адренэргических нейронов, а отрицательные эмоции так же как страх и тревога - с недостатком возбуждения норадренэргических нейронов.
Лимбическая система участвует в организации ориентировочно-исследовательского поведения. Так, в гиппокампе обнаружены нейроны “новизны”, меняющие свою импульсную активность при появлении новых раздражителей. Гиппокамп играет существенную роль в поддержании внутренней среды организма, участвует в процессах обучения и памяти.
Следовательно, лимбическая система организует процессы саморегуляции поведения, эмоции, мотивации и памяти (рис.42).
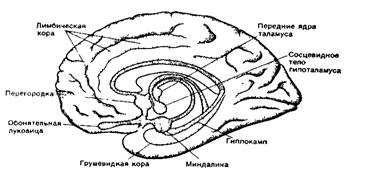
Рис. 42. Лимбическая система
Автономная нервная система
Автономная (вегетативная) нервная система обеспечивает регуляцию внутренних органов, усиливая или ослабляя их деятельность, осуществляет адаптивно-трофическую функцию, регулирует уровень метаболизма (обмен веществ) в органах и тканях (рис.43, 44).
1 - симпатический ствол; 2 - шейно-грудной (звездчатый) узел; 3 – средний шейный узел; 4 - верхний шейный узел; 5 - внутренняя сонная артерия; 6 - чревное сплетение; 7 - верхнее брыжечное сплетение; 8 - нижнее брыжечное сплетение
Рис. 43. Симпатическая часть вегетативной нервной системы,

III- глазодвигательный нерв; YII - лицевой нерв; IX - языкоглоточный нерв; X - блуждающий нерв.
1 - ресничный узел; 2 - крылонебный узел; 3 - ушной узел; 4 - подниж-нечелюстной узел; 5 - подъязычный узел; 6 - парасимпатическое крестцовое ядро; 7 - экстрамуральный тазовый узел.
Рис. 44. Парасимпатическая часть вегетативной нервной системы.
Автономная нервная система включает отделы как центральной, так и периферической нервной системы. В отличие от соматической, в автономной нервной системе эфферентная часть состоит из двух нейронов: преганглионарного и постганглионарного. Преганглионарные нейроны расположены в центральной нервной системе. Постганглионарные нейроны участвуют в образовании автономных ганглиев.
В автономной нервной системе различают симпатический и парасимпатический отделы.
В симпатическом отделе преганглионарные нейроны находятся в боковых рогах спинного мозга. Аксоны этих клеток (преганглионарные волокна) подходят к симпатическим ганглиям нервной системы, расположенным по обе стороны позвоночника в виде симпатической нервной цепочки.
В симпатических ганглиях располагаются постганглионарные нейроны. Их аксоны выходят в составе спинномозговых нервов и образуют синапсы на гладких мышцах внутренних органов, желез, стенок сосудов, кожи и других органов.
В парасимпатической нервной системе преганглионарные нейроны располагаются в ядрах ствола мозга. Аксоны преганглионарных нейронов идут в составе глазодвигательного, лицевого, языкоглоточного и блуждающего нервов. Кроме того, преганглионарные нейроны находятся также в крестцовом отделе спинного мозга. Их аксоны идут к прямой кишке, мочевому пузырю, к стенкам сосудов, снабжающих кровью органы, расположенные в области таза. Преганглионарные волокна образуют синапсы на постганглионарных нейронах парасимпатических ганглиев, расположенных вблизи эффектора или внутри него (в последнем случае парасимпатический ганглий называют интрамуральным).
Все отделы автономной нервной системы подчинены высшим отделам центральной нервной системы.
Отмечен функциональный антагонизм симпатической и парасимпатической нервной системы, что имеет большое приспособительное значение (см. табл. 1).
РАЗДЕЛ IV. РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ
Нервная система начинает развиваться на 3-ей неделе внутриутробного развития из эктодермы (наружного зародышевого листка).
На дорсальной (спинной) стороне зародыша происходит утолщение эктодермы. Это формируется нервная пластинка. Затем нервная пластинка изгибается вглубь зародыша и образуется нервная бороздка. Края нервной бороздки смыкаются, формируя нервную трубку. Длинная полая нервная трубка, лежащая сначала на поверхности эктодермы, отделяется от нее и погружается внутрь, под эктодерму. Нервная трубка расширяется на переднем конце, из которого позднее формируется головной мозг. Остальная часть нервной трубки преобразуется в головной мозг (рис. 45).
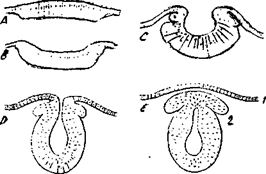
Рис. 45. Стадии эмбриогенеза нервной системы в поперечном схематическом разрезе, а - медуллярная пластинка; b и с - медуллярная бороздка; d и е- мозговая трубка. 1 - роговой листок (эпидермис); 2 - ганглиозный валик.
Из клеток, мигрирующих из боковых стенок нервной трубки, закладываются два нервных гребня - нервные тяжи. В дальнейшем из нервных тяжей образуются спинальные и автономные ганглии и шванновские клетки, которые формируют миелиновые оболочки нервных волокон. Кроме того, клетки нервного гребня участвуют в образовании мягкой и паутинной оболочек мозга. Во внутреннем слове нервной трубки происходит усиленное деление клеток. Эти клетки дифференцируются на 2 типа: нейробласты (предшественники нейронов) и спонгиобласты (предшественники глиальных клеток). Одновременно с делением клеток головной конец нервной трубки подразделяется на три отдела - первичные мозговые пузыри. Соответственно они называются передний (I пузырь), средний (II пузырь) и задний (III пузырь) мозг. В последующем развитии мозг делится на конечный (большие полушария) и промежуточный мозг. Средний мозг сохраняется как единое целое, а задний мозг делится на два отдела, включающих мозжечок с мостом и продолговатый мозг. Это 5-ти пузырная стадия развития мозга ( рис.46,47).
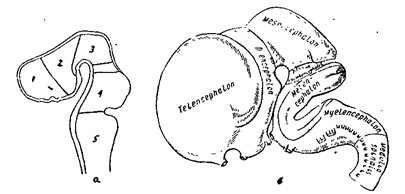
а - пять мозговых путей: 1 - первый пузырь (конечный мозг); 2 - второй пузырь (промежуточный мозг); 3 - третий пузырь (средний мозг); 4- четвертый пузырь (продолговатый мозг); между третьим и четвертым пузырем - перешеек; б - развитие головного мозга (по Р. Синельникову).
Рис. 46. Развитие головного мозга (схема)
|
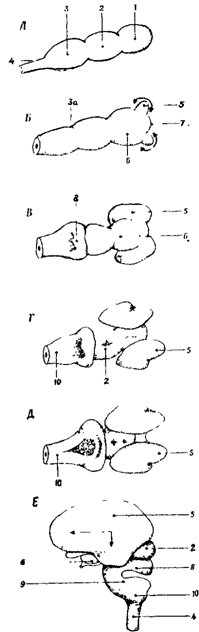
А - формирование первичных пузырей (до 4-й недели эмбрионального развития). Б - Е - формирование вторичных пузырей. Б,В - конец 4-й недели; Г - шестая неделя; Д - 8-9-я недели, завершающиеся формированием основных отделов мозга (Е) - к 14 неделе.
3а — перешек ромбовидного мозга; 7 конечная пластинка.
Стадия А: 1, 2, 3 — первичные мозговые пузыри
1 - передний мозг,
2 - средний мозг,
3 - задний мозг.
Стадия Б: передний мозг делится на полушария и базальные ядра (5) и промежуточный мозг (6)
Стадия В: ромбовидный мозг (3а) подразделяется на задний мозг, включающий в себя мозжечок (8), мост (9) стадия Е и продолговатый мозг (10) стадия Е
Стадия Е: образуется спинной мозг (4)
Рис. 47. Развивающийся мозг.
Образование нервных пузырей сопровождается появлением изгибов, обусловленных разной скоростью созревания частей нервной трубки. К 4-ой неделе внутриутробного развития формируются теменной и затылочный изгибы, а в течение 5-ой недели - мостовой изгиб. К моменту рождения сохраняется только изгиб мозгового ствола почти под прямым углом в области соединения среднего и промежуточного мозга (рис 48).
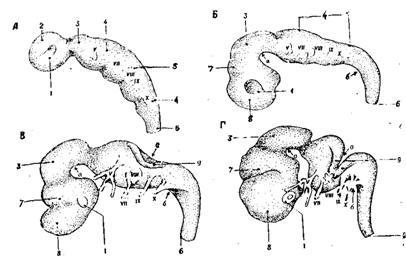
Вид сбоку, иллюстрирующий изгибы в среднемозговой (А), шейной (Б) областях мозга, а также в области моста (В).
1 - глазной пузырь, 2 - передний мозг, 3 - средний мозг; 4 - задний мозг; 5 - слуховой пузырек; 6 - спинной мозг; 7 - промежуточный мозг; 8 - конечный мозг; 9 - ромбическая губа. Римскими цифрами обозначены места отхождения черепно-мозговых нервов.
Рис. 48. Развивающийся мозг (с 3-й по 7-ю неделю развития).
В начале поверхность больших полушарий гладкая, Первыми на 11-12 неделе внутриутробного развития закладывается боковая борозда (Сильвиева), затем центральная (Ролландова) борозда. Довольно быстро происходит закладка борозд в пределах долей полушарий, за счет образования борозд и извилин увеличивается площадь коры ( рис.49).
|
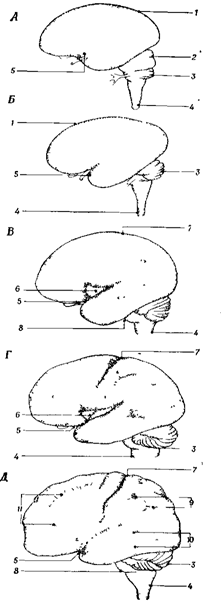
Рис. 49. Вид сбоку на развивающиеся полушария головного мозга.
А- 11-я неделя. Б- 16_ 17 недели. В- 24-26 недели. Г- 32-34 недели. Д - новорожденный. Показано образование боковой щели (5), центральной борозды (7) и других борозд и извилин.
I - конечный мозг; 2 - средний мозг; 3 - мозжечок; 4 - продолговатый мозг; 7 - центральная борозда; 8 - мост; 9 - борозды теменной области; 10 - борозды затылочной области;
II - борозды лобной области.
Нейробласты путем миграции образуют скопления - ядра, формирующие серое вещество спинного мозга, а в стволе мозга - некоторые ядра черепно-мозговых нервов.
Сомы нейробластов имеют округлую форму. Развитие нейрона проявляется в появлении, росте и ветвлении отростков (рис. 50). На мембране нейрона образуется небольшое короткое выпячивание на месте будущего аксона - конус роста. Аксон вытягивается и по нему доставляются питательные вещества к конусу роста. В начале развития у нейрона образуется большее число отростков по сравнению с конечным числом отростков зрелого нейрона. Часть отростков втягивается в сому нейрона, а оставшиеся растут в сторону других нейронов, с которыми они образуют синапсы.

Рис. 50. Развитие веретенообразной клетки в онтогенезе человека. Две последние зарисовки показывают разницу в строении этих клеток у ребенка в возрасте двух лет и взрослого человека
В спинном мозге аксоны имеют небольшую длину и формируют межсегментарные связи. Более длинные проекционные волокна формируются позднее. Несколько позже аксона начинается рост дендритов. Все разветвления каждого дендрита образуются из одного ствола. Количество ветвей и длина дендритов не завершается во внутриутробном периоде.
Увеличение массы мозга в пренатальный период происходит в основном за счет увеличения количества нейронов и количества глиальных клеток.
Развитие коры связано с образование клеточных слоев (в коре мозжечка - три слоя, а в коре полушарий большого мозга - шесть слоев).
В формировании корковых слоев большую роль играют так называемые глиальные клетки. Эти клетки принимают радиальное положение и образуют два вертикально ориентированных длинных отростка. По отросткам этих радиальных глиальных клеток происходит миграция нейронов. Вначале образуются более поверхностные слои коры. Глиальные клетки принимают также участи в образовании миелиновой оболочки. Иногда одна глиальная клетка участвует в образовании миелиновых оболочек нескольких аксонов.
В таблице 2 отражены основные этапы развития нервной системы зародыша и плода.
Таблица 2.
Основные этапы развития нервной системы в пренатальный период.
|
Возраст зародыша (недели) |
Развитие нервной системы |
| 2,5 | Намечается нервная бороздка |
| 3.5 | Образуется нервная трубка и нервные тяжи |
| 4 | Образуются 3 мозговых пузыря; формируются нервы и ганглии |
| 5 | Формируются 5 мозговых пузырей |
| 6 | Намечаются мозговые оболочки |
| 7 | Полушария мозга достигают большого размера |
| 8 | В коре появляются типичные нейроны |
| 10 | Формируется внутренняя структура спинного мозга |
| 12 | Формируются общие структурные черты головного мозга; начинается дифференцировка клеток нейроглии |
| 16 | Различимы доли головного мозга |
| 20-40 | Начинается миелинизация спинного мозга (20 неделя), появляются слои коры (25 недель), формируются борозды и извилины (28-30 недель), начинается миелинизация головного мозга (36-40 недель) |
Таким образом, развитие головного мозга в пренатальный период происходит непрерывно и параллельно, однако характеризуется гетерохронией: скорость роста и развития филогенетически более древних образований больше, чем филогенетически более молодых образований.
Ведущую роль в росте и развитии нервной системы во внутриутробный период играют генетические факторы. Вес мозга новорожденного в среднем составляет примерно 350 г.
Морфо-функциональное созревание нервной системы продолжается в постнатальный период. Уже к концу первого года жизни вес мозга достигает 1000 г, тогда как у взрослого человека вес мозга составляет в среднем - 1400 г. Следовательно, основное прибавление массы мозга приходится на первый год жизни ребенка.
Увеличение массы мозга в постнатальный период происходит в основном за счет увеличения количества глиальных клеток. Количество нейронов не увеличивается, так как они теряют способность делиться уже в пренатальном периоде. Общая плотность нейронов (количество клеток в единице объема) уменьшается за счет роста сомы и отростков. У дендритов увеличивается количество ветвлений.
В постнатальном периоде продолжается также миелинизация нервных волокон как в центральной нервной системе, так и нервных волокон, входящих в состав периферических нервов (черепно-мозговых и спинномозговых.).
Рост спинномозговых нервов связан с развитием опорно-двигательного аппарата и формированием нервно-мышечных синапсов, а рост черепно-мозговых нервов с созреванием органов чувств.
Таким образом, если в пренатальном периоде развитие нервной системы происходит под контролем генотипа и практически не зависит от влияния внешней окружающей среды, то в постанатальном периоде все большую роль приобретают внешние стимулы. Раздражение рецепторов вызывает афферентные потоки импульсов, которые стимулируют морфо-функциональное созревание мозга.
Под влиянием афферентных импульсов на дендритах корковых нейронов образуются шипики - выросты, представляющие собой особые постсинаптические мембраны. Чем больше шипиков, тем больше синапсов и тем большее участие принимает нейрон в обработке информации.
На протяжении всего постнатального онтогенеза вплоть до пубертатного периоде также как и в пренатальный период развитие мозга происходит гетерохронно. Так, окончательное созревание спинного мозга происходит раньше, чем головного мозга. Развитие стволовых и подкорковых структур, раньше, чем корковых, рост и развитие возбудительных нейронов обгоняет рост и развитие тормозных нейронов. Это общие биологические закономерности роста и развития нервной системы.
Морфологическое созревание нервной системы коррелирует с особенностями ее функционирования на каждом этапе онтогенеза. Так, более раннее дифференцирование возбудительных нейронов по сравнению с тормозными нейронами обеспечивает преобладание мышечного тонуса сгибателей над тонусом разгибателей. Руки и ноги плода находятся в согнутом положении - это обуславливает позу, обеспечивающую минимальный объем, благодаря чему плод занимает меньшее место в матке.
Совершенствование координации движений, связанных с формированием нервных волокон, происходит на протяжении всего дошкольного и школьного периодов, что проявляется в последовательном освоении позы сидения, стояния, ходьбы, письма и т.д.
Увеличение скорости движений обуславливается в основном процессами миелинизации периферических нервных волокон и увеличения скорости проведения возбуждения нервных импульсов.
Более раннее созревание подкорковых структур по сравнению с корковыми, многие из которых входят в состав лимбической структуры, обуславливают особенности эмоционального развития детей (большая интенсивность эмоций, неумение их сдерживать связана с незрелостью коры и ее слабым тормозным влиянием).
В пожилом и старческом возрасте происходят анатомические и гистологические изменения мозга. Часто происходит атрофия коры лобной и верхней теменной долей. Борозды становятся шире, желудочки мозга увеличиваются, объем белого вещества уменьшается. Происходит утолщение мозговых оболочек.
С возрастом нейроны уменьшаются в размерах, при этом количество ядер в клетках может увеличиться. В нейронах уменьшается также содержание РНК, необходимой для синтеза белков и ферментов. Это ухудшает трофические функции нейронов. Высказывается предположение, что такие нейроны быстрее утомляются.
В старческом возрасте нарушается также кровоснабжение мозга, стенки кровеносных сосудов утолщаются и на них откладываются холестериновые бляшки (атеросклероз). Это также ухудшает деятельность нервной системы.
ЛИТЕРАТУРА
Атлас “Нервная система человека”. Сост. В.М. Асташев. М., 1997.
Блюм Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум и поведение. М.: Мир, 1988.
Борзяк Э.И., Бочаров В.Я., Сапина М.Р. Анатомия человека. - М.: Медицина, 1993. Т.2. 2-е изд., перераб. и доп.
Загорская В.Н., Попова Н.П. Анатомия нервной системы. Программа курса. МОСУ, М., 1995.
Кишш-Сентаготаи. Анатомический атлас человеческого тела. - Будапешт, 1972. 45-е изд. Т. 3.
Курепина М.М., Воккен Г.Г. Анатомия человека. - М.: Просвещение, 1997. Атлас. Изд.2-е.
Крылова Н.В., Искренко И.А. Мозг и проводящие пути (Анатомия человека в схемах и рисунках). М.: Изд-во Российского университета дружбы народов, 1998.
Мозг. Пер. с англ. Под ред. Симонова П.В. - М.: Мир, 1982.
Морфология человека. Под ред. Б.А. Никитюка, В.П. Чтецова. - М.: Изд-во МГУ, 1990. С. 252-290.
Привес М.Г., Лысенков Н.К., Бушкович В.И. Анатомия человека. - Л.: Медицина, 1968. С. 573-731.
Савельев С.В. Стереоскопический атлас мозга человека. М., 1996.
Сапин М.Р., Билич Г.Л. Анатомия человека. - М.: Высшая школа, 1989.
Синельников Р.Д. Атлас анатомии человека. - М.: Медицина, 1996. 6-е изд. Т. 4.
Шаде Дж., Форд Д. Основы неврологии. - М.: Мир, 1982.
[1] Ткань - это совокупность клеток и межклеточного вещества, сходных по строению, происхождению и выполняемым функциям.
2 Некоторые анатомы продолговатый мозг не включают в задний мозг, а выделяют его в качестве самостоятельного отдела.
Перепечатка материалов без ссылки на наш сайт запрещена